Introduction
Chemical Constituents
Pachymic acid (PA)
Triterpenoids, polysaccharides, and amino acids
Pharmacological Activities
Neuroprotection
Anti-Inflammatory Effects
Anticancer effects
Anti-Ischemia/Reperfusion(I/R) Effects
Hypoglycemic effects
Other effects
Conclusion
Introduction
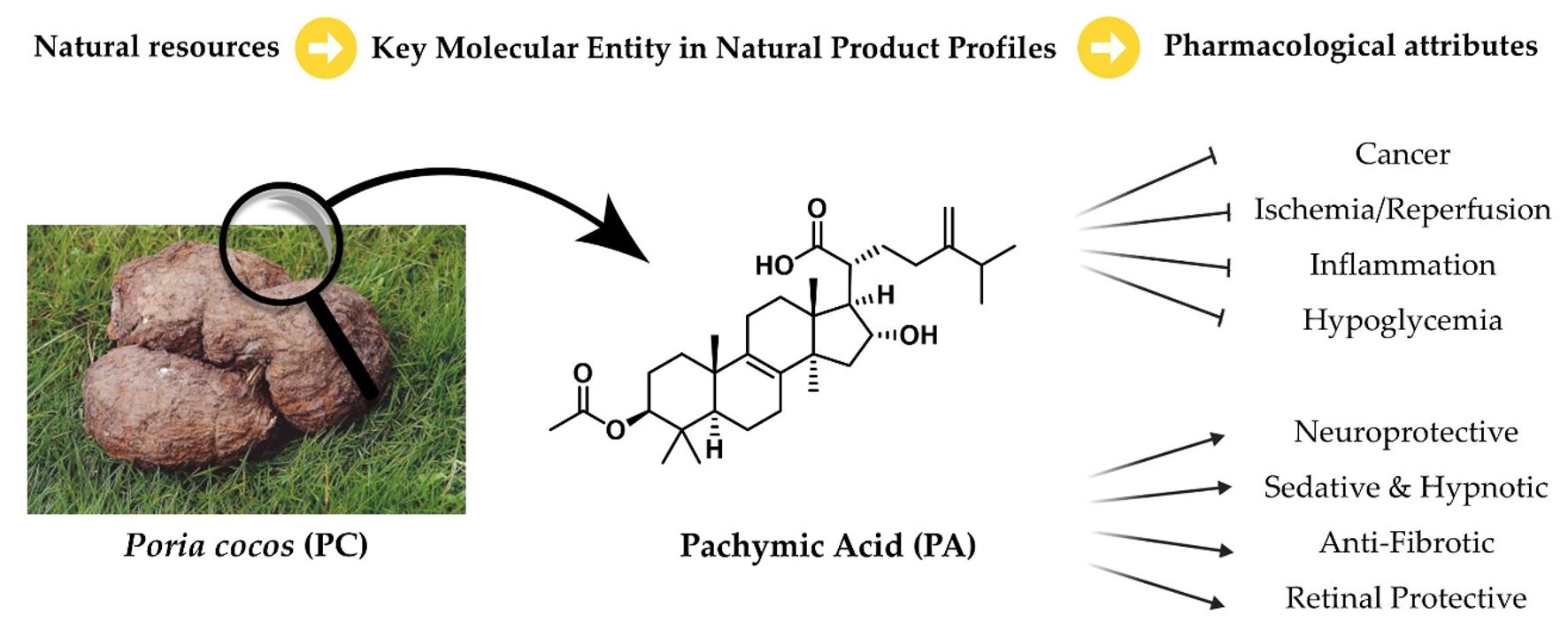
Poria cocos (PC) is a saprophytic fungus that grows naturally around the roots of various Pinus species, and its sclerotium has been widely used for thousands of years in traditional Chinese medicine as well as in functional foods. In Korean medicine, PC is valued for its diuretic, sedative, and spleen-fortifying properties and has been used for the treatment of phlegm retention, edema, kidney disorders, and anxiety. Historical records indicate that PC has been consumed as a food and medicinal fungus for more than two millennia, and modern pharmacological studies have further demonstrated its broad spectrum of biological benefits, including antitumor, hypoglycemic, anti-inflammatory, antimicrobial, and immunomodulatory effects. To establish the precise biological identity of the source fungus, the taxonomic classification of Wolfiporia cocos (F. A. Wolf) is presented in Table 1 (Ryvarden and Gilbertson 1984). Details can be verified through ‘Species Korea’, a database managed by the National Institute of Biological Resources, Republic of Korea. PC contains a range of bioactive constituents, such as triterpenoids, polysaccharides, and amino acids. Among these, pachymic acid (PA), a lanostane-type triterpenoid, has been identified as a key bioactive molecule responsible for multiple physiological functions, with documented causal roles in neuroprotection (Lin et al. 2009) anti-inflammatory (Kim et al. 2013) and anticancer effects (Ma et al. 2015), ischemia/reperfusion (I/R) protection (Pang et al. 2020), and blood glucose regulation (Mehan et al. 2011). In 2022, a research group in China published a comprehensive review detailing the biosynthetic pathways, pharmacokinetic properties, metabolic processes, and molecular mechanisms underlying PA’s biological activities, making an important contribution that helped renew interest in PA as a promising candidate for natural-product-based drug development (Wei et al. 2022). Building on this momentum, we sought to trace the recent progression of pharmacological research on PA. In this review, we summarize the biological characteristics and pharmacological potential of PA reported in the scientific literature from 2022 to the present. For completeness and comparative context, some studies published before 2022 are also included.
Table 1.
Taxonomic classification of Poria cocos (F. A. Wolf)*.
| Appearance | Division | Name |
 | Kingdom | Fungi |
| Phylum | Basidiomycota | |
| Class | Agaricomycetes | |
| Order | Polyporales | |
| Family | Polyporaceae | |
| Genus | Wolfiporia | |
| Species | Wolfiporia cocos (F. A. Wolf) |
Chemical Constituents
Pachymic acid (PA)
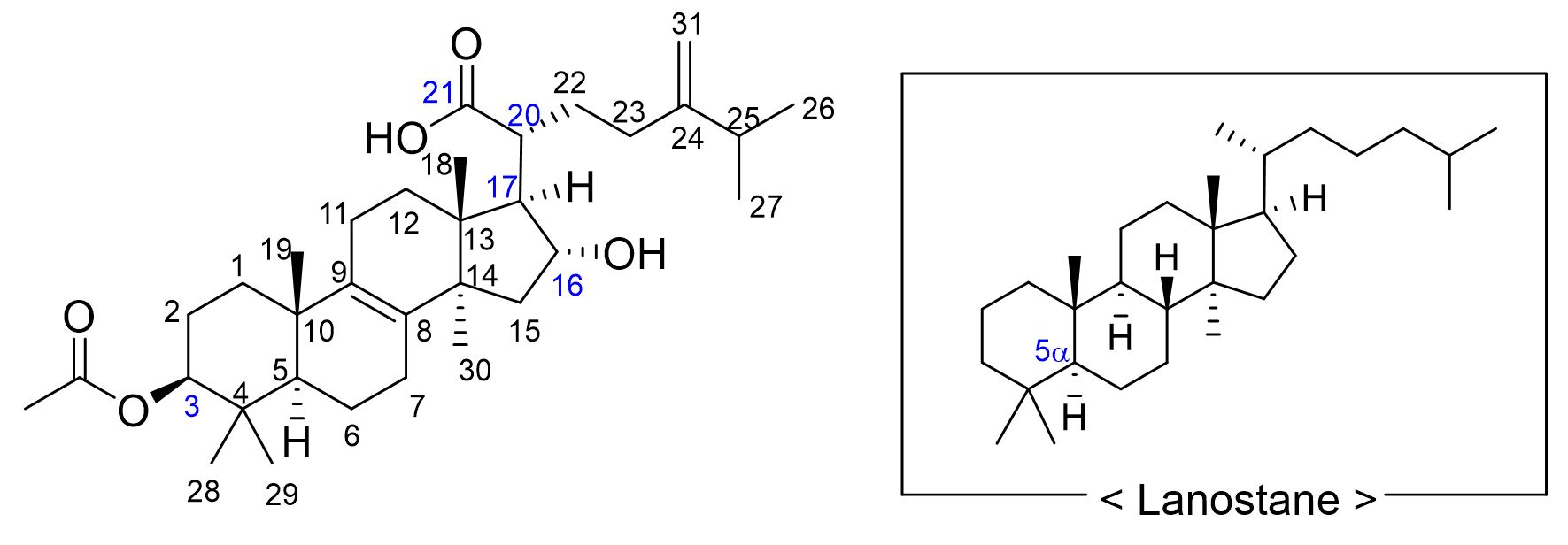
PA was first identified in the sclerotium of PC and its structural properties as a lanostane-type triterpenoid were further elucidated in subsequent studies (Shibata et al. 1958). PA is a naturally occurring white powder that is highly insoluble in water but is known for its low toxicity and minimal adverse effects. Structurally, PA is a tetracyclic triterpenoid with a 5α-lanostane core (Fig. 1). It carries an acetoxy group at C3 (from the lanosterol backbone), an α-hydroxyl group at C16, and a carboxylic acid at C21. The biosynthetic pathway constrains C17 and C20 to be the R configuration. The embedding and spatial arrangement of these functional groups influence their acidity, solubility in polar solvents, and electrostatic interactions, thereby affecting the biological activity.
Triterpenoids, polysaccharides, and amino acids
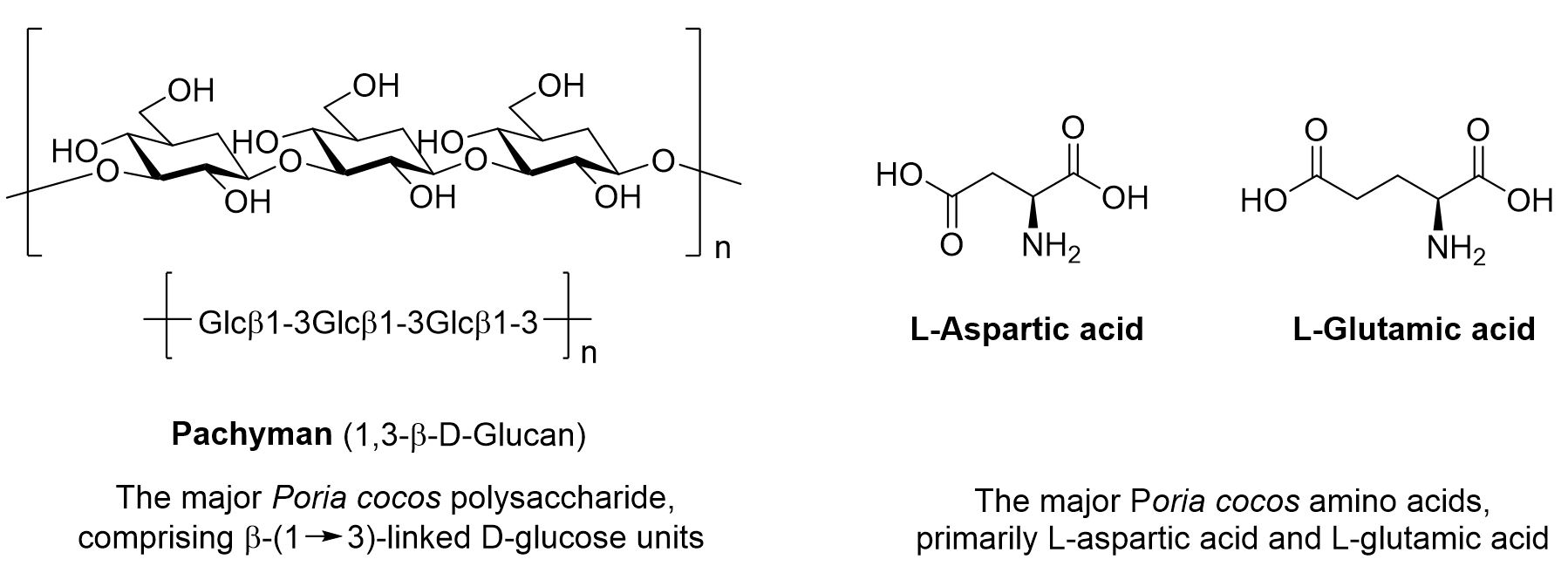
In addition to PA, PC contains various bioactive compounds such as triterpenoids, polysaccharides, and amino acids, each possessing distinct physiological properties and pharmacological activities (Fig. 2). These constituents are found in numerous natural products and medicinal plants, and their biological effects vary depending on their structural characteristics and content levels. Triterpenoids are polysaccharide-based compounds that typically have a C30 structure. Polysaccharides are high molecular weight polymers composed of multiple monosaccharide units and are known to enhance the immune system and regulate various physiological responses. Amino acids play essential roles in protein synthesis, neurotransmission, and metabolic processes. These components work in concert to support diverse physiological functions and contribute to the prevention and treatment of various diseases.
Pharmacological Activities
Neuroprotection
Among the diverse pharmacological attributes of PA, neuroprotection is arguably its most significant therapeutic dimension. In light of a recent surge in landmark reports confirming its potent impact on the central nervous system, this section prioritizes PA’s role in preserving neurological integrity as a focal point of its bioactivity. By bridging systemic recovery with cellular resilience, PA stands at the forefront of current research into PC-derived interventions for brain health. These consolidated findings are summarized in Table 2.
Table 2.
Neuroprotective effects of PA
| Entity | Models | Mechanism / Details | References |
| 3.1.1 | Neuron-like rat pheochromocytoma PC12 cells | Atractylenolide II, III, levistolid A, and ferulic acid work together to exhibit a much stronger neuroprotective effect than a single ingredient and prevent nerve cell necrosis and death by inhibiting lipid peroxidation, preventing energy depletion, and suppressing nerve cell excitation. | Lin et al. 2009 |
| 3.1.2 | SH-SY5Y cells | PA regulates IL-1β, CXCL8, CCL7, and LRP1 in KEGG pathways such as the NF-κB signaling pathway and IL-17 signaling pathway, preventing Fe2+ induced apoptotic cell death. | Hu et al. 2024a |
Preventive and Therapeutic Effects of FDA Extract in Cerebral I/R Injury
In mice subjected to repeated cerebral I/R, high levels of neurospecific enolase (NSE) leakage and widespread brain infarction were observed 24 hours after injury. However, mice that received oral pretreatment with either FDA or FDA-CO₂ extract exhibited a significant reduction in infarct area and a notable decrease in circulating NSE levels compared with sham-operated controls (p < 0.05). This study provides further evidence that the FDA extract can help prevent and mitigate I/R-induced nerve damage (Lin et al. 2009).
Reduction of Iron-Induced Reactive Oxygen Species (ROS) and Enhancement of Mitochondrial Function
Accumulating evidence has further elucidated the cytoprotective mechanisms of PA, specifically concerning its role in maintaining oxidative homeostasis and mitochondrial stability. Treatment with PA effectively attenuates the accumulation of iron-induced reactive oxygen species (ROS) and facilitates the restoration of mitochondrial membrane potential, thereby preempting the initiation of apoptotic cascades. These cellular responses suggest that the neuroprotective efficacy of PA is fundamentally anchored in its ability to modulate the neuroinflammatory-oxidative axis. By neutralizing oxidative stress-related injury, PA reinforces the metabolic resilience of neurons, offering a robust defense against degenerative pathological conditions (Hu et al. 2024)
Anti-Inflammatory Effects
Inflammation represents a pivotal physiological and pathological axis implicated in the pathogenesis of various systemic conditions, including malignancy, metabolic disorders, and cardiovascular diseases. Extensive research has substantiated that PA possesses potent anti-inflammatory properties, with several key regulatory pathways identified to date. PA acts as a multi-target modulator that suppresses pro-inflammatory cascades, thereby mitigating tissue damage across diverse disease models. The specific anti-inflammatory actions and the associated molecular mechanisms of PA are comprehensively summarized in Table 3.
Table 3.
Anti-inflammatory effects of PA
| Entity | Models | Mechanism / Details | References |
| 3.2.1 | MC-3T3 E1 cells induced with AH Plus | Restore AH Plus damaged cell viability and ALP activity, suppress secretion of NO, TNF‑α and IL‑1β, reduce ROS formation and NF‑κB translocation | Kim et al. 2013 |
| 3.2.2 | Rats were anesthetized with 0.5% sodium pentobarbital intraperitoneally and LPS was administered intranasally to induce acute lung injury. | PA improved the histopathological alterations in lung tissues in model rats with pneumonia. PA inhibited the expression levels of inflammatory cytokines in rat serum and suppressed cell apoptosis from rat lung tissue. Mechanically, PA inhibited inflammation and cell apoptosis via NF-κB and MAPK pathways. | Gui et al. 2021 |
| 3.2.3 | Articular cartilage was harvested from the knee and femoral head of C57BL/6 mice. | PA delays the development of OA by promoting the expression of SIRT6 and inhibiting the NF-κB pathway in IL-1β stimulated chondrocytes. | Wu et al. 2023 |
| 3.2.4 | Raw 264.7 cells | Treatment with p-Cymene and PA resulted in a decrease in the levels of proinflammatory cytokines, namely TNF-α, IL-1β, and IL-6, while concurrently increasing the level of IL-10. | Gu et al. 2024a |
Anti-Inflammatory Effects of PA and Its Potential in Preventing Oral Inflammatory Disorders
The pharmacological efficacy of PA extends to the protection of bone-forming cells within inflammatory microenvironments. Research has demonstrated that PA can effectively alleviate osteocyte damage induced by exogenous irritants, such as the dental sealer AH Plus. At the cellular level, PA treatment significantly suppresses the expression of pro-inflammatory mediators in MC-3T3 E1 cells while concurrently upregulating endogenous antioxidant signaling pathways. These findings suggest that PA attenuates material-induced bone damage by neutralizing oxidative stress and dampening the inflammatory response, thereby promoting the structural and functional integrity of osteogenic tissues (Kim et al. 2013)
Protection Mechanism of PA Against Renal and Pulmonary Injury: Regulation of MAPK, AMPK, and PI3K/Akt Pathways
The anti-inflammatory and cytoprotective properties of PA have also been substantiated in models of acute respiratory distress, such as pneumonia. Studies utilizing in vivo pneumonia models have demonstrated that PA administration significantly attenuates inflammatory infiltration and suppresses the expression of apoptosis-related markers within lung tissue. This pulmonary protection is primarily mediated through the orchestrated regulation of the NF-κB and MAPK signaling pathways, which are pivotal in the progression of systemic inflammation. By dampening these signaling cascades, PA effectively reduces pathological cell death and mitigates tissue-level injury, suggesting its potential as a therapeutic intervention for inflammatory lung diseases (Gui et al. 2021)
Anti-Inflammatory Effects of PA and the Protection Mechanism Against Osteoarthritis via SIRT6
The therapeutic potential of PA has been substantiated in experimental models of chronic kidney disease (CKD). Research indicates that the administration of PA effectively ameliorates histopathological renal damage and leads to a significant reduction in biomarkers associated with chronic inflammation and interstitial fibrosis. These renoprotective outcomes are primarily attributed to the ability of PA to modulate the Wnt/β-catenin-angiotensin axis, a critical signaling pathway in the progression of renal deterioration. By regulating this axis, PA mitigates the fibrotic cascade and preserves functional renal integrity, suggesting its role as a promising pharmacological candidate for CKD intervention (Wu et al. 2023)
Therapeutic Potential of p-Cymene and PA in Inflammatory Bowel Disease (IBD) via Regulation of Signaling Pathways
The anti-inflammatory efficacy of Pachymic acid (PA) has been further extended to the context of pharmacological resistance. Experimental evaluations in adalimumab-resistant cell lines have demonstrated that treatment with PA, often in conjunction with compounds such as p-cymene, significantly attenuates inflammatory responses. This effect is primarily mediated through the suppression of the PI3K/Akt/mTOR signaling pathway, which leads to a marked reduction in downstream pro-inflammatory biomarkers. These findings position PA as a potential therapeutic candidate for managing inflammation in cells that have developed resistance to conventional biologics. However, it should be noted that current evidence is predominantly based on in vitro data, and further in vivo studies are necessitated to validate these preliminary therapeutic outcomes (Gu et al. 2024a)
Anticancer effects
The World Health Organization estimates that annual cancer cases are expected to increase substantially, from 18 million in 2018 to approximately 29.5 million by 2040 (Wu et al. 2023), highlighting the urgent need for more effective and less toxic anticancer agents. Natural products are an important source of such agents, with approximately 30–40% of clinically used anticancer drugs estimated to originate from plant-derived compounds. Among the major bioactive constituents of PC, PA has demonstrated notable anticancer potential. Relevant studies and their progression are summarized in Table 4.
Table 4.
Anticancer effects of PA
| Entity | Models | Mechanism / Details | References |
| 3.3.1 | NCl-H23, NCl-H460 lung cancer cells | PA induces apoptosis at least in part by activating the JNK and ER stress apoptotic pathways, as shown by TUNEL staining in tumor tissues. | Mehan et al. 2011; Ma et al. 2015 |
| 3.3.2 | LNCaP and DU145 prostate cancer cells | PA reduced AA production in a manner similar to 4-bromophenacyl bromide (4BPB), a nonspecific pharmacological inhibitor of PLA2. | Gapter et al. 2005 |
| 3.3.3 | BALB/c nude mice that have been subcutaneously injected with breast cancer cells MDA-MB-231 | PA downregulated the apoptotic regulator Bcl-2, increased the expression of proapoptotic protein Bax, and promoted the release of cytochrome c and the activation of cleaved caspase-3, -9 and caspase-8 via mitochondria-mediated and death receptor-mediated signaling pathways. | Jiang and Fan 2020 |
| 3.3.4 | HGC-27 and GES1 cells | PA reduced the viability of HGC-27 cells, halted the cell cycle, enhanced cell apoptosis, regulated the PI3K/Akt signaling pathway to promote apoptosis, and restricted cell proliferation. | Du et al. 2024 |
Lung Cancer Cell Growth Inhibition and Apoptosis Induction by PA via JNK and Endoplasmic Reticulum (ER) Stress Pathways
Mehan et al. reviewed the role of the stress-activated c-Jun N-terminal kinases (JNKs) in various neurodegenerative diseases, including Alzheimer’s disease. They found that JNK signaling plays a key role in oxidative stress, inflammation, and apoptosis, suggesting its importance as a potential target for future treatment of neurodegenerative diseases (Mehan et al. 2011). In 2015, Ma et al. examined whether PA could induce apoptosis in lung cancer cell lines. Their findings demonstrated that PA treatment increased intracellular ROS levels and activated both the JNK and ER stress pathways, ultimately leading to apoptosis. These results indicate that PA can inhibit lung cancer cell growth by promoting apoptosis through ROS-dependent activation of JNK and ER stress pathways, highlighting its potential as an anticancer agent (Ma et al. 2015)
Inhibition of Prostate Cancer Cell Growth by PA and Its Time-Dependent Effects
Gapter et al. investigated whether PA extracted from PC could induce apoptosis in human prostate cancer cells. They discovered that PA suppressed the growth of both androgen-insensitive DU145 and LNCaP prostate cancer cell lines in a dose- and time-dependent manner, with more pronounced inhibition observed in DU145 cells. PA treatment increased p21 expression without causing cell-cycle arrest and activated mitochondrial apoptosis, as evidenced by reduced Bad phosphorylation, increased Bcl-2 phosphorylation, and activation of caspase-9 and caspase-3. The study also found that Akt signaling components were downregulated, indicating that PA may promote apoptosis by inhibiting prostaglandin synthesis and Akt activity (Gapter et al. 2005)
Effects of PC and PA on Suppressing Breast Cancer Cell Survival and Reducing Adverse Reactions
In 2020, Jiang and Fan investigated the anticancer activity of an ethanol extract of PC using both in vitro and in vivo methods. Following chromatographic purification, they identified PA as a major active compound that demonstrated strong cytotoxicity against MDA-MB-231 breast cancer cells (IC₅₀ = 2.13 ± 0.24 µg/mL) while showing no appreciable effect on normal mammary epithelial MCF-10A cells, indicating a high degree of selectivity. In BALB/c nude mice injected with MDA-MB-231 cells, PC treatment significantly reduced the final tumor weight (0.51 ± 0.12 g) compared with untreated mice. Notably, PC produced fewer adverse effects on major organ function and muscular strength relative to cisplatin. The authors found that PC in vitro inhibited MDA-MB-231 growth in a dose-dependent manner and induced apoptosis alongside G0/G1 cell-cycle arrest. Specifically, treatment downregulated cyclin D1, cyclin E, CDK2, and CDK4, while upregulating p53and p21. Additional evidence of apoptosis included decreased apoptotic regulator Bcl-2, increased Bax, cytochrome c release, and activation of caspases-3, -9, and -8, indicating engagement of both mitochondrial and death receptor-mediated pathways (Jiang and Fan 2020)
Network Pharmacology Analysis of PA in Gastric Cancer (GC) Treatment and Regulation of the PI3K/Akt Pathway
In 2024, Du et al. investigated the therapeutic mechanisms of PA in GC treatment using a combination of network pharmacology and experimental validation. Through multiple bioinformatics databases, the authors identified potential targets related to both GC and PA, with SRC, MAPK1, PIK3R1, HSP90AA1, and PTPN11 noted as key targets. STRING and KEGG.GO analyses suggested that the PI3K/Akt pathway is likely the main mechanism involved, and molecular docking was performed to support these predicted interactions. In vitro experiments using HGC-27 GC cells demonstrated that PA reduced cell viability, induced cell-cycle arrest, and promoted apoptosis. Notably, expression of PI3K/Akt pathway proteins was suppressed, while apoptosis-related proteins showed characteristic changes, including increased Bax, decreased Bcl-2, and activation of caspases. In vivo studies and TCGA-based sequencing analyses confirmed that PA targets PDGFRB to inhibit the PI3K/Akt pathway, thereby inducing ferroptosis and suppressing GC progression. Overexpression of PDGFRB weakened the antitumor effects of PA, while ferroptosis-associated markers, such as increased MDA, Fe²⁺, and ROS levels, along with reduced GSH, became prominent. Together, these findings support PA as a potential GC treatment candidate and provide a stronger basis for its potential use in clinical settings (Du et al. 2024)
Anti-Ischemia/Reperfusion(I/R) Effects
PA has demonstrated remarkable efficacy in mitigating I/R injury, a complex pathological process characterized by acute metabolic collapse followed by a surge in oxidative stress upon reoxygenation. The therapeutic hallmark of PA in this context lies in its ability to modulate tissue-specific signaling cascades—including the PI3K/Akt, NRF2, and AMPK pathways—to suppress diverse forms of programmed cell death, such as apoptosis and ferroptosis. By stabilizing cellular homeostasis and preserving microvascular integrity across multiple organ systems, PA functions as a versatile protective agent against the systemic cascades of I/R-induced damage. The multi-organ protective effects and their underlying molecular targets are comprehensively summarized in Table 5.
Table 5.
Anti- I/R Effects PA
| Entity | Models | Mechanism / Details | References |
| 3.4.1 | Healthy male SD rats | Increase cerebral blood flow, reduce infarct volume and brain water content, and decrease neuronal damage and neuronal apoptosis via upregulation of p-PTEN, p-PDK1, p-Akt and p-BAD and downregulation of cleaved caspase protein expression | Pang et al. 2020 |
| 3.4.2 | C57BL/6 mice of renal I/R injury | PA activates the NRF2 signaling pathway, leading to the upregulation of antioxidant proteins such as GPX4, xCT, and HO-1. This promotes glutathione (GSH) biosynthesis and the elimination of lipid peroxides, thereby inhibiting ferroptosis and protecting the kidney from acute injury. | Jiang et al. 2021 |
| 3.4.3 | Male wild-type (WT) mice and AMPK knockout (−/−) mice (C57BL/6J background) | PA inhibited ferroptosis of cardiomyocytes through activating of the AMPK pathway, thereby alleviating myocardial I/R injury in mice. | Liu et al. 2024 |
Reduction of Cerebral I/R Injury Through the PI3K/Akt Pathway
In 2020, Pang et al. examined the neuroprotective effects of PA in rats with cerebral I/R injury, focusing on the PI3K/Akt signaling pathway. The authors found that PA treatment improved several indicators of brain recovery, including increased cerebral blood flow, reduced infarct size, alleviated brain edema, and improved neurological scores. It also reduced post-ischemic neuronal damage and apoptosis, as shown by Nissl and TUNEL staining. Western blot results demonstrated that PA enhanced the expression of key PI3K/Akt pathway proteins, including p-PTEN (Ser380), p-PDK1 (Ser241), p-Akt (Ser473), pc-Raf (Ser259), and p-BAD (Ser136), while reducing levels of cleaved caspase. Notably, the signal pathway inhibitor LY294002 abolished the protective effects of PA, supporting the conclusion that PI3K/Akt activation is central to its mechanism. These findings indicate that PA exerts significant neuroprotective effects against cerebral I/R injury and neuronal apoptosis, likely through activation of the PI3K/Akt signaling pathway (Pang et al. 2020).
Kidney Protection in I/R-Induced Acute Kidney Injury (AKI)
Jiang et al. investigated whether PA could protect the kidneys in an I/R-induced AKI model by suppressing ferroptosis. The authors found that rats treated with PA showed lower serum creatinine and BUN levels, less structural kidney damage, and reduced mitochondrial injury. They also reported that PA inhibited ferroptosis, as evidenced by increased expression of GPX4, SLC7A11 (xCT), and HO-1, along with decreased levels of MDA and COX-2. These effects were associated with activation of the NRF2 signaling pathway, suggesting that PA may help regulate ferroptosis and could serve as a viable treatment option for AKI (Jiang et al. 2021)
Mechanisms Underlying the Protective Effects of PA Against Myocardial I/R Injury
Liu et al. examined whether PA could mitigate myocardial I/R injury by suppressing ferroptosis in cardiomyocytes. The authors observed that PA-treated mice showed less myocardial tissue damage, had decreased Fe²⁺ accumulation and lipid peroxidation, and increased cardiomyocyte viability. Molecular mechanism analyses revealed that PA activated the AMPK signaling pathway, which subsequently upregulated GPX4 and SLC7A11 (xCT), thereby contributing to the suppression of ferroptosis. These findings suggest that PA may protect the heart during myocardial I/R injury by regulating ferroptosis through the AMPK pathway activation (Liu et al. 2024)
Hypoglycemic effects
PA acts as a multifaceted modulator of glucose homeostasis, exhibiting both direct pharmacodynamic effects on insulin sensitivity and significant pharmacokinetic influences on co-administered antidiabetic agents. Beyond its primary role in reducing blood glucose levels through PPAR-γ-independent pathways, PA exerts a systemic impact on the metabolic environment by inhibiting specific cytochrome P450 enzymes. This dual characteristic underscore PA’s potential as a functional antidiabetic ingredient while necessitating careful clinical consideration regarding potential drug-drug interactions (DDIs). The comprehensive antihyperglycemic profiles and metabolic interactions of PA are summarized in Table 6.
Table 6.
hypoglycemic effects of PA
| Entity | Models | Mechanism / Details | References |
| 3.5.1 | Male C57BL/KsJ-db/db mice, which possess a point mutation of the leptin receptor, and male C57BL/6J mice | PC extract and its triterpenes reduce postprandial blood glucose levels in db/db mice via enhanced insulin sensitivity, irrespective of PPAR-γ. | Li et al. 2011 |
| 3.5.2 | Male SD rats | PA increased systemic exposure and decreased the clearance of repaglinide, suggesting an inhibitory effect of PA on repaglinide metabolism. | Gu et al. 2024b |
Antidiabetic Effects and Mechanisms of PC and Its Active Triterpenes
Li et al. evaluated whether a crude extract of PC and its isolated triterpenes exert hypoglycemic effects in a type 2 diabetes mouse model. The authors found that administration of the crude extract at doses ≥50 mg/kg significantly reduced both fasting and postprandial blood glucose levels in db/db mice. Bioactivity-guided fractionation identified dehydrotumulosic acid, dehydrotrametenolic acid, and PA as the major active constituents, among which dehydrotumulosic acid exhibited the strongest glucose-lowering effect, while PA also demonstrated significant activity. In streptozotocin (STZ)-treated mice, both the crude extract and the triterpenes improved insulin sensitivity, resulting in lower blood glucose levels, although they did not directly activate the PPAR-γ pathway. Overall, the study suggests that PC-derived triterpenes can improve insulin sensitivity and help control postprandial glucose independently of PPAR-γ, supporting their potential use as functional antidiabetic ingredients (Li et al. 2011)
Drug–Drug Interaction Between PA and Repaglinide and the Underlying Metabolic Inhibition Mechanism
Gu et al. investigated how PA influences the metabolism of repaglinide, an antidiabetic medication. The authors examined changes in repaglinide plasma levels in rats treated with PA and found that rats receiving both PA and repaglinide showed higher repaglinide concentrations than the control group. This effect was attributed to PA’s inhibition of CYP3A4, an enzyme essential for repaglinide metabolism, which slowed its clearance. These findings suggest that using PA and repaglinide together may raise repaglinide plasma levels and increase the risk of adverse effects such as hypoglycemia, highlighting the need for caution when the two are co-administered (Gu et al. 2024b)
Other effects
Beyond its previously detailed bioactivities, PA demonstrates a versatile pharmacological spectrum by modulating diverse physiological systems, including neurotransmission, protein folding homeostasis, and sensory preservation. PA acts as a multi-target regulator that restores systemic balance—specifically by enhancing GABAergic inhibitory signaling, mitigating fibroproliferative cascades through the suppression of endoplasmic reticulum (ER) stress, and safeguarding retinal integrity against age-related oxidative damage. This broad-spectrum efficacy highlights PA’s potential as a systemic homeostatic stabilizer capable of addressing complex, multi-factorial pathologies. These multifaceted effects are summarized in Table 7.
Table 7.
Other effects of PA
| Entity | Models | Mechanism / Details | References |
| 3.6.1 | ICR male mice | In culture models, protein levels of the α- and β-subunits of the GABAa receptor were overexpressed, but treatment of cells with PA resulted in a decrease in the γ-subunit of the rat GABAa receptor. | Shah et al. 2014 |
| 3.6.2 | BLM-induced Specific Pathogen Free (SPF) SD rats | PA may ameliorate BLM-induced pulmonary fibrosis and histopathological damage in rats through inhibiting ER stress and improving mitochondrial function. | Li et al. 2024 |
| 3.6.3 | A retinal oxidative damage model induced by injecting sodium iodate (SI) into the tail vein of mice | PA could activate Nrf2, thereby upregulating the expression of the downstream antioxidant gene HO-1 protein, enhancing the ability of cells to clear ROS, maintaining the balance of the redox state in cells, inhibiting the oxidative stress response, and protecting cells from oxidative stress damage. | Qin et al. 2024 |
Sedative and Hypnotic Effects of PA: Mechanistic Actions via the GABA-ergic System
Shah et al. investigated the effects of PA on pentobarbital-induced sleep behavior in mice. The authors reported that PA administration shortened sleep onset latency and significantly prolonged sleep duration, with these effects becoming more pronounced when PA was co-administered with pentobarbital. In addition, PA increased the expression of GAD65/67—key enzymes involved in GABA synthesis—and upregulated the α and β subunits of the GABAa receptor, while downregulating the γ subunit. These findings suggest that PA may exert sedative and hypnotic effects on the central nervous system by regulating the GABAa-ergic system (Shah et al. 2014)
Anti-Fibrotic Effects of PA and Underlying Mechanisms
Li et al. examined whether PA could reduce lung damage in a bleomycin-induced pulmonary fibrosis model by suppressing ER stress. The authors administered PA orally at 10, 20, and 40 mg/kg for 14 d and found notable decreases in lung edema and fibrosis, along with improvements in histopathological changes. Levels of hydroxyproline and TGF-β1 were significantly decreased, and the expression of fibrosis-related proteins (collagen I, α-SMA, and fibronectin) was also reduced. Further mechanism analyses showed increases in antioxidant indicators (SOD, CAT, ATP) and decreases in MDA and ROS. Notably, the expression of ER stress-related proteins (GRP78, CHOP, Caspase-9, and ATF4) was significantly downregulated. These findings suggest that PA may help reduce oxidative stress and mitigate bleomycin-induced pulmonary fibrosis by suppressing ER stress and improving mitochondrial function (Li et al. 2024)
Retinal Protective Effects of PA via the Nrf2/HO-1 Pathway and Its Potential in Preventing Age-Related Macular Degeneration
Qin et al. investigated the protective effects and underlying mechanisms of PA in oxidative stress retinal injury models induced by sodium iodate (SI) or hydrogen peroxide. In vitro, they used ARPE-19 cells exposed to H₂O₂ to establish an oxidative stress model and found that PA pretreatment restored cell viability and increased Bcl-2 expression while reducing apoptosis, ROS production, and the expression of Bax and caspase-3. In vivo, the authors found that PA administration substantially reduced NaIO₃-induced retinal atrophy and apoptosis. OCT imaging, HE, and TUNEL staining results further confirmed improvements in retinal thickness and reductions in cell death. Collectively, these results suggest that PA may help protect against age-related macular degeneration by reducing oxidative stress levels (Qin et al. 2024)
Conclusion
This study reviewed the therapeutic potential of PA, a major triterpenoid derived from PC, and its effects across a wide range of diseases (Fig. 3). Evidence from prior studies and recent experimental findings indicates that PA exhibits diverse biological and pharmacological activities, including neuroprotective, anti-inflammatory, antioxidant, anticancer, antidiabetic, and organ-protective effects. These activities are mediated through multiple signaling pathways—such as JNK, NF-κB, SIRT6, PI3K/Akt, MAPK, Nrf2/HO-1, and AMPK—allowing PA to suppress inflammation and oxidative stress and to mitigate the progression of disease through diverse molecular mechanisms involved in cell survival, differentiation, and death. In addition, PA has demonstrated efficacy across a variety of disease models, including neurological disorders, osteoarthritis, IBD, diabetes, and lung cancer, and in some cases, synergistic effects were observed when combined with conventional treatments. In organ injury models, PA contributed to protecting tissues and supported recovery by regulating inflammatory mediators and preserving tissue structure. Taken together, these findings suggest that PA, with its low toxicity and broad range of bioactivities, represents a promising therapeutic candidate for the treatment of multiple diseases. Further studies are needed to deepen our understanding of its mechanisms, along with evaluations of toxicity, pharmacokinetics, and potential clinical applications.