

Introduction
Seagrass degradation and loss
Restoration of seagrass
Genetic diversity and restoration
Conclusion
Introduction
Seagrasses are flowering marine plants that evolved from terrestrial plants around 149 million years ago (Silva et al. 2021). Seagrasses harbor rich diversity, with 72 species reported worldwide (Short et al. 2011). Unlike marine macroalgae, seagrasses have true roots, stems, fruits and leaves. Their shoots and rhizomes can form dense meadows in shallow coastal waters, serving as an important ecosystem to maintain coastal health by supporting food security, biodiversity, habitat and nursery grounds for fish and invertebrates, carbon sequestration, reducing coastal erosion, and improving water quality (do Amaral Camara et al. 2023). Unfortunately, this fragile ecosystem has been threatened globally. There has been an overall declining trend across all seven bioregions amounting to about 5,602 km² (19.1% of the surveyed meadow area) since 1880 (Dunic et al. 2021). In Southeast Asia, seagrass meadows was estimated at 3,670 km² (Sudo and Nakaoka 2020), forming the most diverse and productive coastal ecosystems in the world. Nevertheless, they have been extensively degraded due to pollution, coastal development, eutrophication, and destructive fishing practices. Urbanization and aquaculture activities have strongly contributed to sedimentation and reduced water quality, thereby further stressing the habitats. Seagrass restoration is one of the most urgent strategies to prevent degradation, by transplanting shoots from healthy meadows. In contrast, seed-based restoration can promote genetic diversity, and hence is suitable for large-scale restoration. Theoretically, populations with higher genetic diversity are more resilient (Connolly et al. 2018), implying that future restoration should include genetic diversity to evaluate success under local environmental and climate changes (Pazzaglia et al. 2021).
In Viet Nam, the updated seagrass distribution is estimated at 15,634 ha (Nguyen-Nhat et al. 2024). Fifteen seagrass species have been recorded, with three dominant species: Enhalus acoroides (Linnaeus f.) Royle, Thalassia hemprichii (Ehrenberg) Ascherson, and Halophila ovalis (R.Brown) J.D.Hooker (Nguyen et al. 2022a). Similar to other countries worldwide, seagrass beds in Viet Nam are experiencing degradation. Major threats include coastal development and habitat destruction, pollution and eutrophication, sedimentation and turbidity, destructive fishing behaviors, and climate change (Orth et al. 2006; Waycott et al. 2009). Small scale restoration for H. ovalis, E. acoroides, and Nanozostera japonica (Ascherson & Graebner) Tomlinson & Posluszny (Fig. 1) have been conducted at Ha Long Bay, Thu Bon estuary (Nguyen 2010), and Thuy Trieu lagoon (Pham 2008). However, genetic diversity of these three species was not considered. In this review, we present the degradation and restoration of seagrass in Viet Nam in comparison with global trends, and highlight the role of genetic diversity in seagrass restoration. In addition, lessons were mentioned from failures and successes of seagrass restoration in the world.

Fig. 1.
Transplantation of Nanozostera japonica from a healthy meadow to a disturbed site for restoration in Nui Thanh, Da Nang. Source: Nguyen (2010).
Seagrass degradation and loss
Despite the ecologically important role, seagrasses are declining globally. Both natural disturbances (e.g. storms, hurricanes, and diseases) and human activities (e.g. dredging, pollution, coastal development, boating, and fishing practices) are the main causes (Strydom et al. 2023). Waycott et al. (2009) reported that approximately 29% of the world’s seagrass beds have been lost or degraded since 1879. Mwikamba et al. (2024) reported annual seagrass loss rates of 0.29–1.59% in Kenya, 0.26–7.2% (1991–2013) in Mozambique, 2% (1972–2010) in Tunisia, and 2.8–38% (1960–2007) in South Africa. Therefore, Africa’s seagrass cover in some areas is declining even faster than the global average annual rate (7%). They also proposed that human-induced factors were the most significant threats to African seagrasses, with 74% of the identified drivers directly linked to human activities. Based on seagrass species and percent cover from a broad area of the Eastern Banks, Australia, Smart et al. (2025) found a decline in seagrass meadows due to changing species composition and reductions in spatial distribution. Another species Posidonia oceanica (Linnaeus) Delile meadows at The Ligurian Sea (northwestern Mediterranean) have lost nearly half of their original beds since the mid-19th century due to intensive coastal development (Montefalcone 2024). In Tampa Bay, Florida, USA, seagrass coverage has declined by about 11,518 acres since 2016 (Beck et al. 2024), which ascribed to alterations in physical water quality associated with climate change. In Australia, seagrass decline is consistent with global patterns, with at least 291,783 ha, equivalent to about 5.5% of the estimated national seagrass coverage lost since the 1930s (Statton et al. 2018).
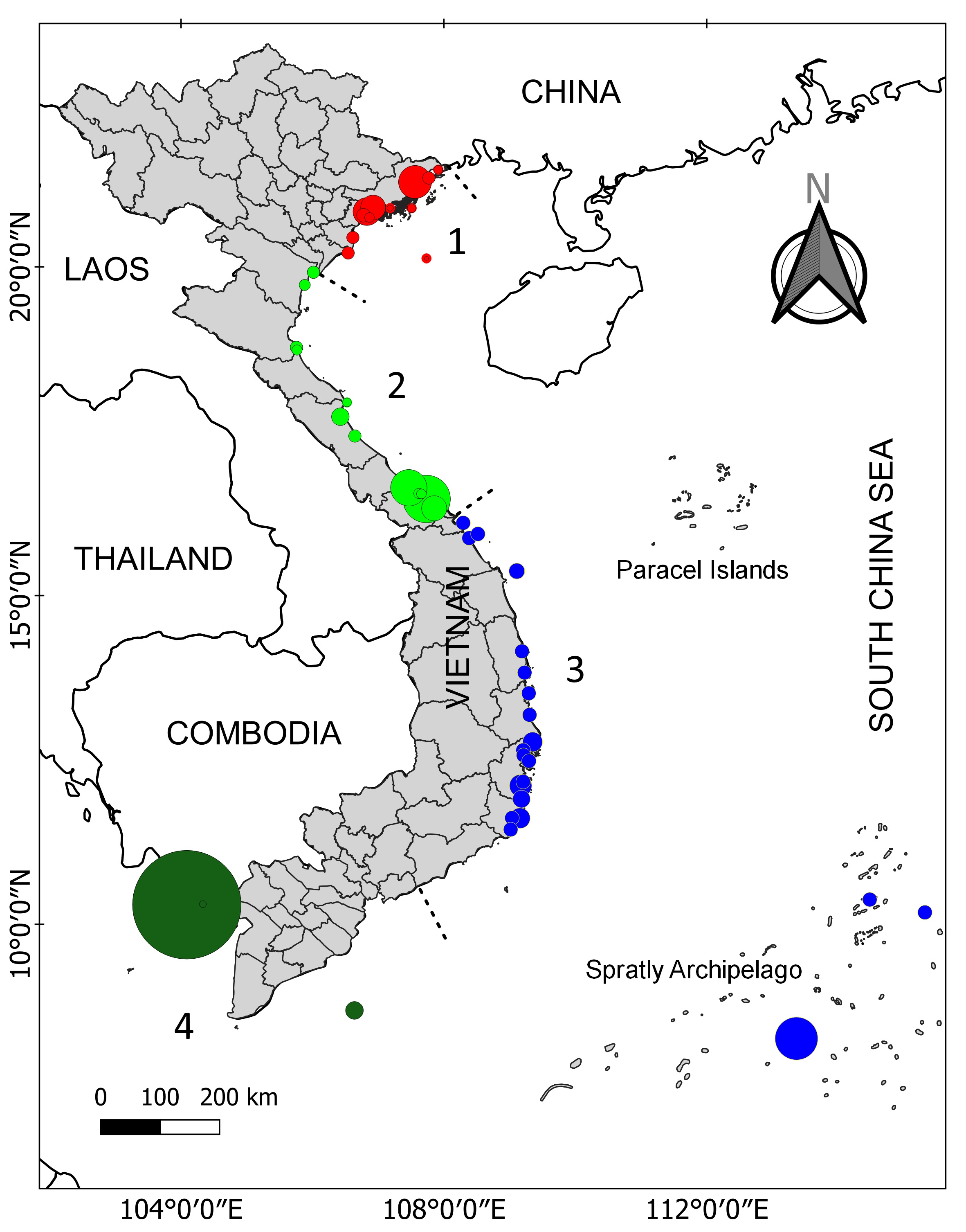
The coastline of Viet Nam is divided into four regions: northeast, north central, south central, and southern Viet Nam (Fig. 2). Seagrasses occur widely along the coast and around offshore islands, with the highest diversity found in the south central and southern regions. However, over the past several decades, the country has experienced a substantial decline in both seagrass extent and health. Nguyen et al. (2004) initially reported seagrass degradation in Khanh Hoa province. Subsequent detailed studies revealed spatial and temporal changes of seagrasses in specific areas. For instance, Vo et al. (2020) reported that 186.2 ha (equivalent to 35.8%) of the original seagrass beds in Van Phong Bay were lost over the past three decades due to various anthropogenic and natural factors. Similarly, Chen et al. (2016) found that the total area of seagrass beds in Cam Ranh Bay declined by approximately 25% (66 ha) from 1996 to 2015, primarily as a result of coastal development and infrastructure expansion. More recently, Nguyen et al. (2021a) estimated that the total coverage of seagrass beds in Southern Viet Nam to be 10,832 ha, with an estimated loss of 2,562 ha (or 19.1%). At the national scale, Trinh and Takeuchi (2019) estimated that seagrass coverage in Viet Nam has declined by about 46.5% (or 13,549 ha) since 1990. Seagrass beds in the West Sea of Viet Nam are undergoing degradation (Fig. 3). Key threats include coastal infrastructure development such as island ring roads and harbor construction as well as small-scale aquaculture activities, particularly fish and Babylonia areolata (sea snail) farming (Nguyen-Nhat et al. 2024).

Restoration of seagrass
Restoration has become an important strategy to reduce the global decline of seagrass ecosystems. Several techniques have been applied, including shoot transplantation and seed-based propagation. Selection of each method strongly depends on site characteristics, species and local environmental conditions (Unsworth et al. 2025). Broadly, restoration methods involve two categories: transplantation and seed-based approaches. Transplantation relocates adult shoots, sods, or plugs from healthy donor beds to degraded beds (Ferretto et al. 2021), whereas seed-based restoration uses the seeds collected from natural populations, which are either sown directly in the field or germinated under controlled conditions (Unsworth et al., 2025). However, survival rates vary widely across species and locations. In Australia, for example, restoration incorporated a wide range of methods such as the transplantation of sprigs, plugs, and seed-based interventions.
These efforts have ranged from small pilot projects to large-cale transplantation trials, incorporating both manual and mechanical planting techniques and employing various anchoring methods, including biodegradable pots, hooks, and pegs. Bastyan and Cambridge (2008) reported that transplanted Amphibolis antarctica (Labillardière) Ascherson exhibited considerable spatial and temporal variability in performance. Recruitment was recorded on all planting bags, and long-term survival (up to 38 months) ranged from 0 to 72 individuals/m². Using seeds sown into hessian, Tanner et al. (2014) demonstrated good recruitment and survival, with older bags (62 months after deployment) showing greater stem lengths than those observed in natural meadows at Grange, Australia. In contrast, restoration attempts involving H. ovalis and Zostera marina Linnaeus showed no evidence of the benefit of transplanting, as survival was virtually zero (Connolly et al. 2016).
For seed-based restoration, Buoy-Deployed Seeding (BDS) and Dispenser Injection Seeding (DIS) models were applied in Australia. The BDS technique involves collecting mature reproductive shoots and placing them in mesh bags attached to floating buoys, which are suspended above the target restoration area (Pickerell et al. 2005). As the shoots release their negatively buoyant seeds, the seeds naturally sink and settle across the intended restoration plot. Although this approach can be applied over relatively large spatial scales, its recruitment remains low, with only 1-6.9% of deployed seeds (Marion and Orth, 2010). DIS is one of the most recently developed techniques in seagrass restoration (Govers et al. 2022). In this method, seagrass seeds are combined with local sediment to form a sediment-seed mixture, which is then injected into the substrate using modified sealant guns. These iterative improvements resulted in an 83-fold increase in field plant densities (from 0.012 to 1.00 plants m-2) and a slight reduction in seed loss between 2015 and 2017, demonstrating the potential of DIS to enhance seagrass establishment in restoration sites. However, these developments have not yet resulted in self-sustaining seagrass populations.
Genetic diversity and restoration
In recently years, understanding genetic diversity and population structure of donor and recipient populations has been proved crucial to guide restoration and conservation strategies (Mijangos et al. 2015). Byeon et al. (2025) indicated that genetic diversity is key to maintaining ecological resilience, as it boosts seagrass population density, supports richer associated faunal communities, and enables quicker recovery after disturbances. The importance of assessing genetic diversity for restoration has been widely emphasized, particularly considering the need to evaluate donor bed diversity before undertaking seagrass transplantation. Recently, several studies stressed the urgent need to understand the relationships among threats, conservation management, and changes in genetic diversity (Moore et al. 2025; Shaw et al. 2025)
In the Eastern Mediterranean Sea, a genetic study of Posidonia oceanica provided valuable information for the restoration. Analyses of twelve microsatellite markers showed that genetic information can identify suitable donor meadows for transplantation and conservation priorities (Litsi-Mizan et al. 2024). Another study at two large-scale sites in Italy: Santa Marinella (Central Tyrrhenian Sea) and Ischia Porto (Southern Tyrrhenian Sea) indicated that shoot density after ten years in transplanted areas reached levels comparable to natural meadows. However, the phenological and lepidochronological traits were still different (Litsi-Mizan et al. 2024). These findings also provide valuable information for P. oceanica restoration. For other species, such as Halophila stipulacea (Forsskål) Ascherson in the Mediterranean, population genetic characteristic has been demonstrated essential for assessing invasion risks and refining conservation strategies for seagrass ecosystems (García-Escudero et al. 2023).
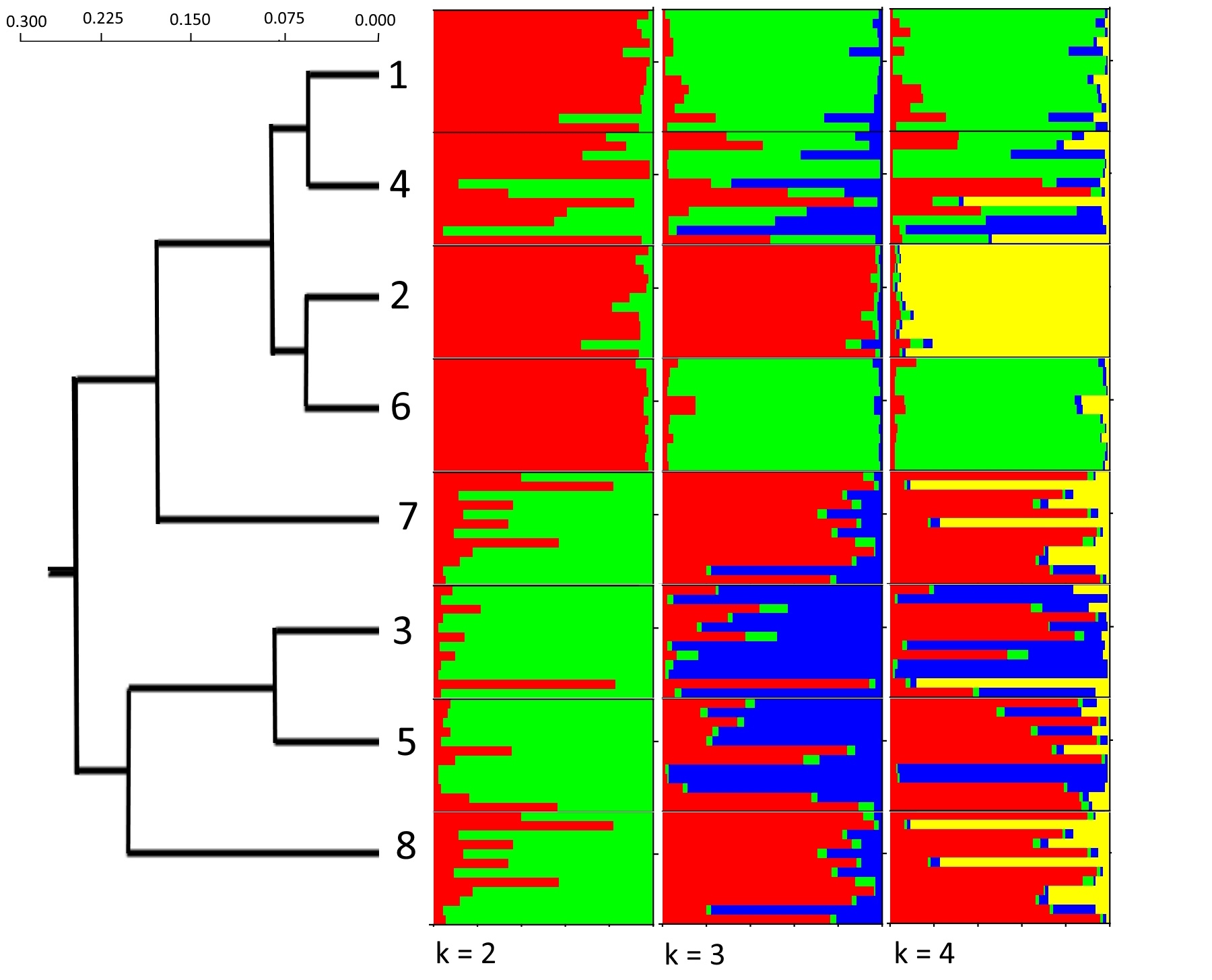
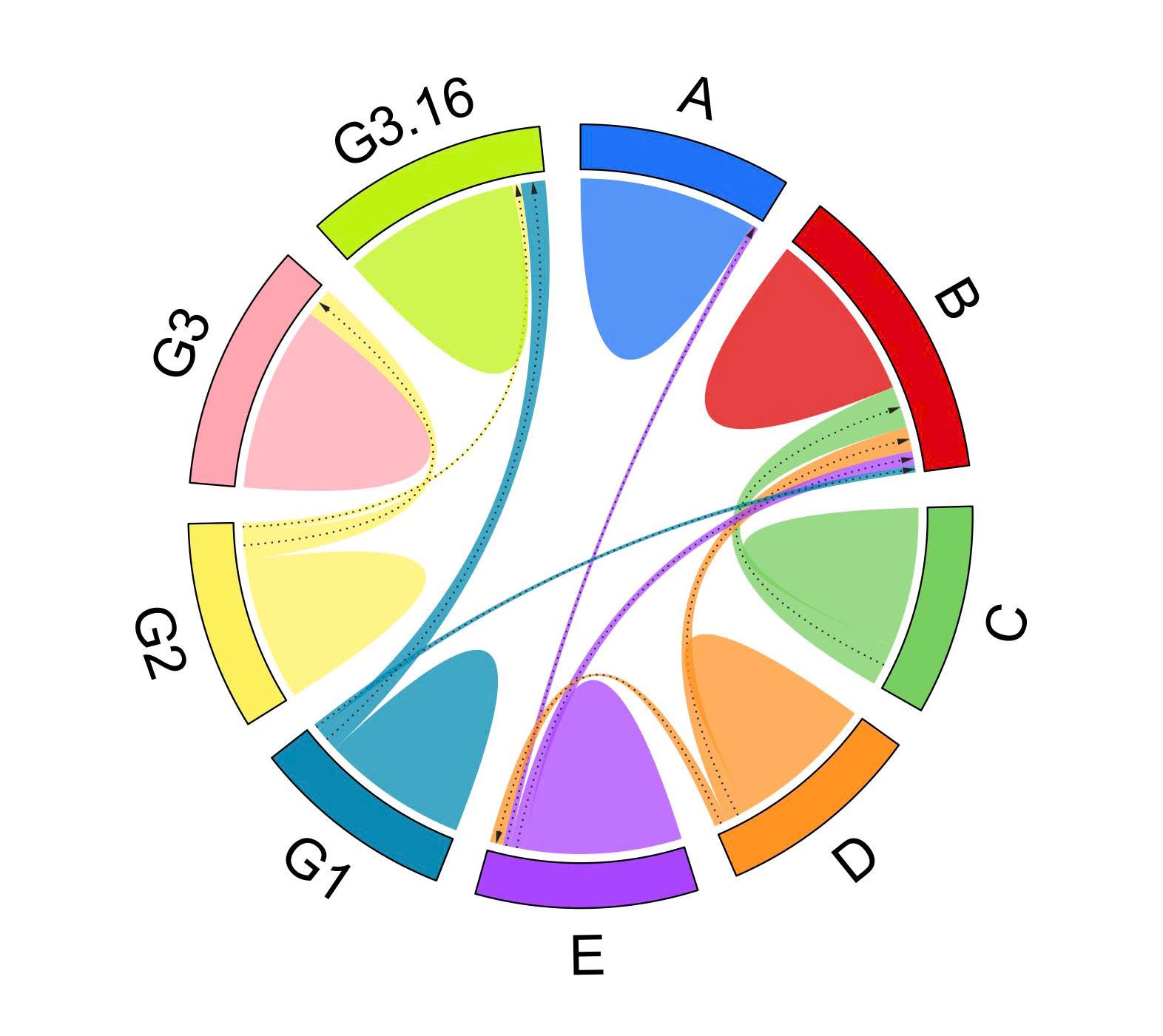
The role of genetic diversity in restoration of seagrass beds in Viet Nam also received increasing attention. Based on ten microsatellite loci, Nguyen et al. (2022b) revealed two genetic groups of T. hemprichii (hard bottom and soft bottom) (Fig. 4), and found that western boundary currents drove gene flow in southern Viet Nam. They also identified four populations with high relative genetic contribution to design conservation priorities. For other species E. acoroides, Nguyen and Papenbrock (2019) found that its meadows along the Khanh Hoa Province coastline exhibited reduced genetic diversity, as well as pronounced genetic differentiation between lagoon sites and open-sea sites. Eight E. acoroides populations along the South-Central Coast of Viet Nam were characterized with clonal richness, genetic diversity, and dispersal within and among populations in four lagoons (Dierick et al. 2021). The strong genetic differentiation among lagoons, suggests that these lagoon systems acted as barriers to gene flow, and large, persistent genets enhanced the resilience of E. acoroides meadows under high disturbance regimes. In addition, the Department of Marine Botany, Institute of Oceanography revealed that three populations of E. acoroides along Khanh Hoa coast may be used as donor sites for transplanting (unpublished data). The gene flows among E. acoroides populations in the Gulf of Thailand and East Sea of Viet Nam were clarified (Fig. 5).
With the support from WWF, a seagrass restoration project is currently being implemented at Cu Lao Cham MPA, Da Nang. Cu Lao Cham is offshore islands, comprising a main island (approximately 15 km²) and seven smaller islands. Small scale seagrass beds occur surrounding these islands. Among the five species found in this area, Halophila decipiens Ostenfeld (Fig. 6) and Halodule pinifolia (Miki) Hartog (Fig. 7) are the two dominant. According to Tin et al. (2020), the coverage of seagrass beds at Cu Lao Cham experienced significant changes between 2003 and 2017. The bed lost 40.17 ha but gained 58.6 ha from 2003 to 2010. In contrast, from 2010 to 2017, seagrass coverage exhibited a loss of 68.76 ha with only 7.27 ha of newly formed beds. They also reported that Halophila ovalis and Nanozostera japonica were the two of five species recorded in this area. However, Cao et al. (2019) did not include Nanozostera japonica and Oceana serrulata (R.Brown) Byng & Christenhusz (former name: Cymodocea serrulata) in the seagrass species list of Cu Lao Cham. Several specimens of Halophila major (Zollinger) Miquel were misidentified as Halophila ovalis due to the overlap of morphological traits between each other. Recently, Nguyen et al. (2021b) noted that H. major mainly occurs in the offshore island, whereas H. ovalis is commonly found in inland lagoons. In addition, the putative hybridization between H. ovalis and H. major has been reported (Liu et al. 2020; Nguyen-Nhat et al. 2024). Therefore, the application of genetic markers is essential for improving our understanding of species identities and genetic diversity of seagrass in Cu Lao Cham MPA, an offshore ecosystem.


For mangroves, recent studies have also emphasized that maintaining genetic diversity is essential for successful restoration because populations with higher genetic diversity exhibit greater adaptive potential, resilience, and long-term ecosystem stability (Wee et al. 2019). In Viet Nam, three decades after the Viet Nam War ended, populations of Avicennia alba were still increasing in the Mekong Delta (Arnaud-Haond et al. 2009). For another species of mangroves, Sonneratia caseolaris, Le and Le (2024) reported high levels of genetic variation both among and within natural populations in Viet Nam, and the authors suggested that conservation of genetic resources is urgently needed for their future use in conservation and reforestation programs. In the South Central Coast of Viet Nam, the genetic diversity data of Rhizophora apiculata provided important recommendations for mangrove breeding and restoration programs (Hong Nguyen et al. 2025). In Brazil, using inter-simple sequence repeat (ISSR) markers for two species including Laguncularia racemosa and Avicennia schaueriana, Granado et al. (2018) indicated that knowledge of genetic monitoring associated with restoration activities can help avoid bottlenecks and other pitfalls. Analyses of genetic diversity and mating systems have been applied for conservation and restoration strategies of Rhizophora apiculata and Avicennia marina in Tielu Harbor Mangrove Nature Reserve, China (Lu et al. 2022). For marine macroalgae, recent studies have highlighted that preserving genetic diversity is critical for successful restoration and long-term resilience (Coleman et al. 2020). Based on genetic baseline data, three populations of the red alga Ahnfeltiopsis pusilla located in the southernmost sites of the Northwest Iberian Peninsula were identified as particularly valuable for conservation because they contain most of the species’ genetic heritage and provide useful insights for the conservation of endangered seaweeds (Couceiro et al. 2011). Kelp forests have experienced localized to regional declines due to multiple stressors, including eutrophication, overgrazing, ocean warming, and marine heatwaves (Carnell and Keough 2019). Understanding genetic structure appears to be a key factor for successful large-scale restoration (Coleman and Wernberg 2018; Wernberg et al. 2018). In support of restoration practices for coastal shrub species (Westringia fruticosa and Acacia suaveolens), Rossetto et al. (2019) suggested that genomic data are an important component in decisions regarding the sourcing of restoration materials to meet specific restoration objectives.
Based on these previous studies, the role of genetic diversity in seagrass restoration can be listed as follows:
ⅰ) Seagrass populations with higher genetic diversity have greater tolerance to environmental changes such as temperature, salinity, turbidity, and nutrients (Hughes and Stachowicz 2004);
ⅱ) Using multiple genotypes in restoration can enhances survival, density, and growth of seagrass. Therefore, it maintains both success and stability of seagrass beds (Pazzaglia et al. 2021);
ⅲ) Due to resistant genotypes, genetic variation can support populations against disease, and help conserve population structure and ecological functions during outbreaks (Ries et al. 2023);
ⅳ) Seagrass beds with high genetic diversity provide stronger services such as carbon storage, sediment stabilization, nutrient cycling, and functions of habitat (Bijak et al. 2023);
ⅴ) Genetic diversity enhances flowering rates, seed production, the success of sexual recruitment, and long-term survival (Alotaibi et al. 2019);
ⅵ) Genetic diversity increases the capacity of populations to adapt to environmental changes such as temperatures and storm, thereby supporting long-term ecosystem stability (Ehlers et al. 2008);
ⅶ) Genetic data can be used to reveal suitable donor beds, ensuring that genotypes are appropriate for sink meadows (Mijangos et al. 2015); and
ⅷ) Introducing genotypes with advantageous traits such as heat or salinity tolerance, can improve restoration outcomes and enhance the resilience of restored seagrass meadows under climate change (Unsworth et al. 2025)
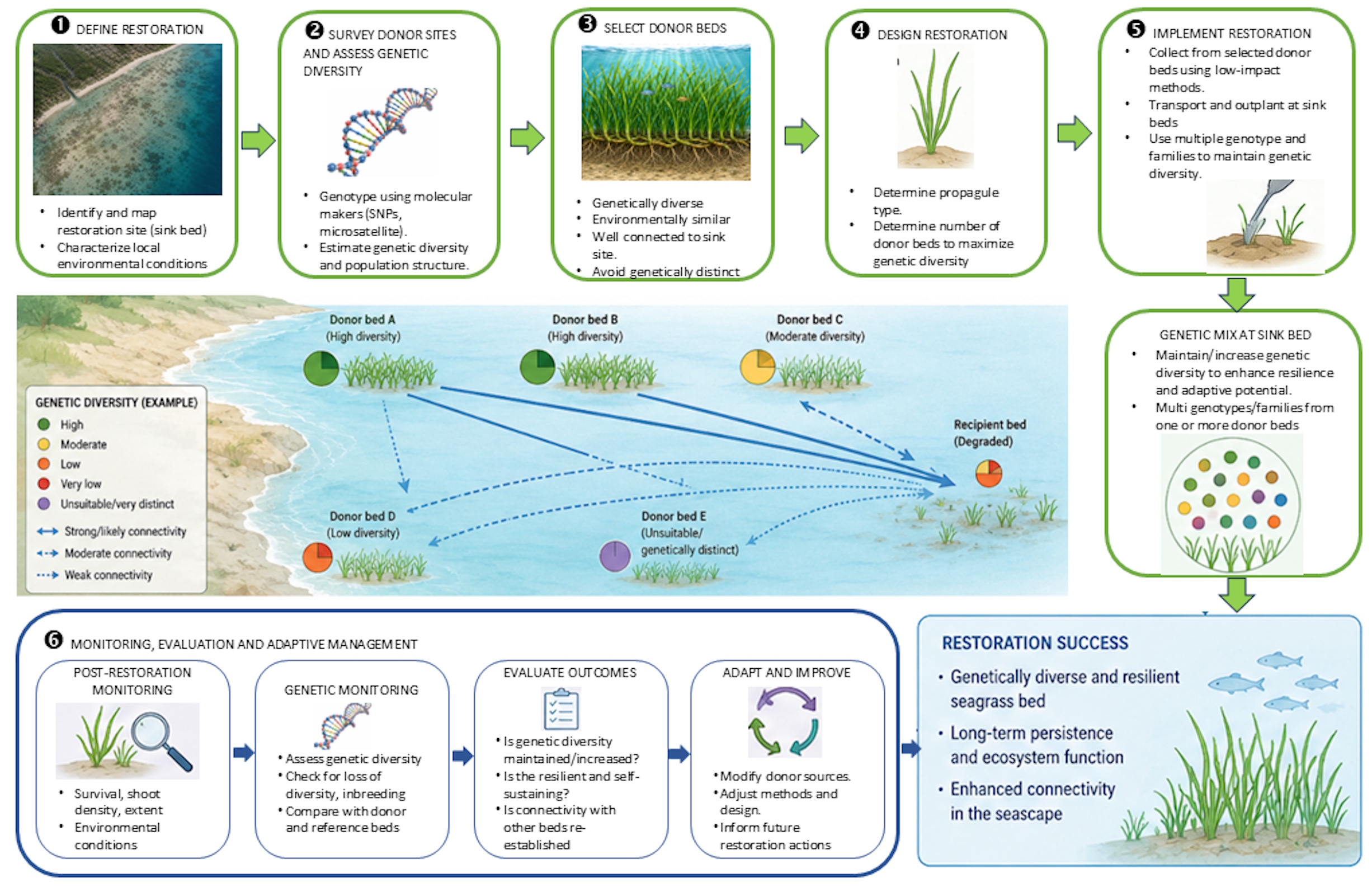
The following diagram illustrates how use genetic diversity information can be used to guide seagrass bed restoration of seagrass beds, including the donor and recipient beds in nature (Fig. 8). We propose six key steps for restoration success of seagrass beds: 1. Define restoration, 2. Survey donor populations and assess genetic diversity, 3. Select donor populations, 4. Design restoration, 5. Implement restoration, and 6. Monitoring, evaluation and adaptive management. Finally, successful seagrass restoration is expected to result in genetically diverse and resilient populations, long-term persistence and ecosystem function, and improved connectivity across the seascape.
Conclusion
This review highlights recent patterns of seagrass degradation, summarizes key restoration techniques, and emphasizes the importance of genetic diversity in supporting successful restoration efforts of seagrass beds. The lessons learned from past restoration initiatives can inform future projects in Viet Nam, where seagrass meadows are increasingly the focus of restoration and conservation activities.
