

Introduction
Porphyrin Biosynthesis Pathways
The common core pathway for the formation of tetrapyrrole structures
Protoporphyrin-dependent pathway
Coproporphyrin-dependent pathway
Siroheme-dependent pathway
Recent Advances in Metabolic Engineering of Various Microorganisms for the Improved Production of Metalloporphyrins
Escherichia coli
Bacillus subtilis
Corynebacterium glutamicum
Saccharomyces cerevisiae
Rhodobacter sphaeroides
Conclusion
Introduction
Metalloporphyrin is a kind of macrocyclic molecule composed of four pyrrole groups with a metal ion in the core (Zhang et al. 2022). It plays an important role in not only biological systems, but also various applications such as photodynamic therapy, dye-sensitized solar cells, catalysts, interface, and food additives (Ko et al. 2024). Most metalloporphyrins used in these applications are chemically synthesized; however, this approach involves lengthy, complex processes with low yields and high energy requirements due to elevated reaction temperatures.
Metalloporphyrins in most living organisms are synthesized via porphyrin metabolism. The biosynthesis of metalloporphyrins is environmentally friendly, as it typically utilizes non-toxic substrates such as carbohydrates under mild reaction conditions (Yu and Zhou 2024). Furthermore, it enables selective synthesis by leveraging the substrate specificity of enzymes. However, metalloporphyrin biosynthesis is often constrained by genetic, enzymatic, and metabolic regulatory mechanisms. To overcome these limitations and improve the biosynthetic yield of metalloporphyrins, extensive research of metabolic engineering in microbes has been conducted in recent years (Ko et al. 2018). Especially, recent studies suggest that not only metabolic engineering using industrial microorganisms but also synthetic biology approaches employing photosynthetic bacteria have made significant contributions to the production of metalloporphyrins.
This review consolidates recent insights into porphyrin biosynthesis pathways, including protoporphyrin-dependent (PPD), coproporphyrin-dependent (CPD), and siroheme-dependent (SHD) pathways, with a focus on enzymatic reactions to help readers better understand the recombinant strain development through metabolic engineering. This review also highlights recent advances in metabolic engineering research that has significantly enhanced metalloporphyrin biosynthesis in microorganisms, such as Escherichia coli, Bacillus subtilis, Corynebacterium glutamicum, Saccharomyces cerevisiae, and Rhodobacter sphaeroides (Fig. 1).
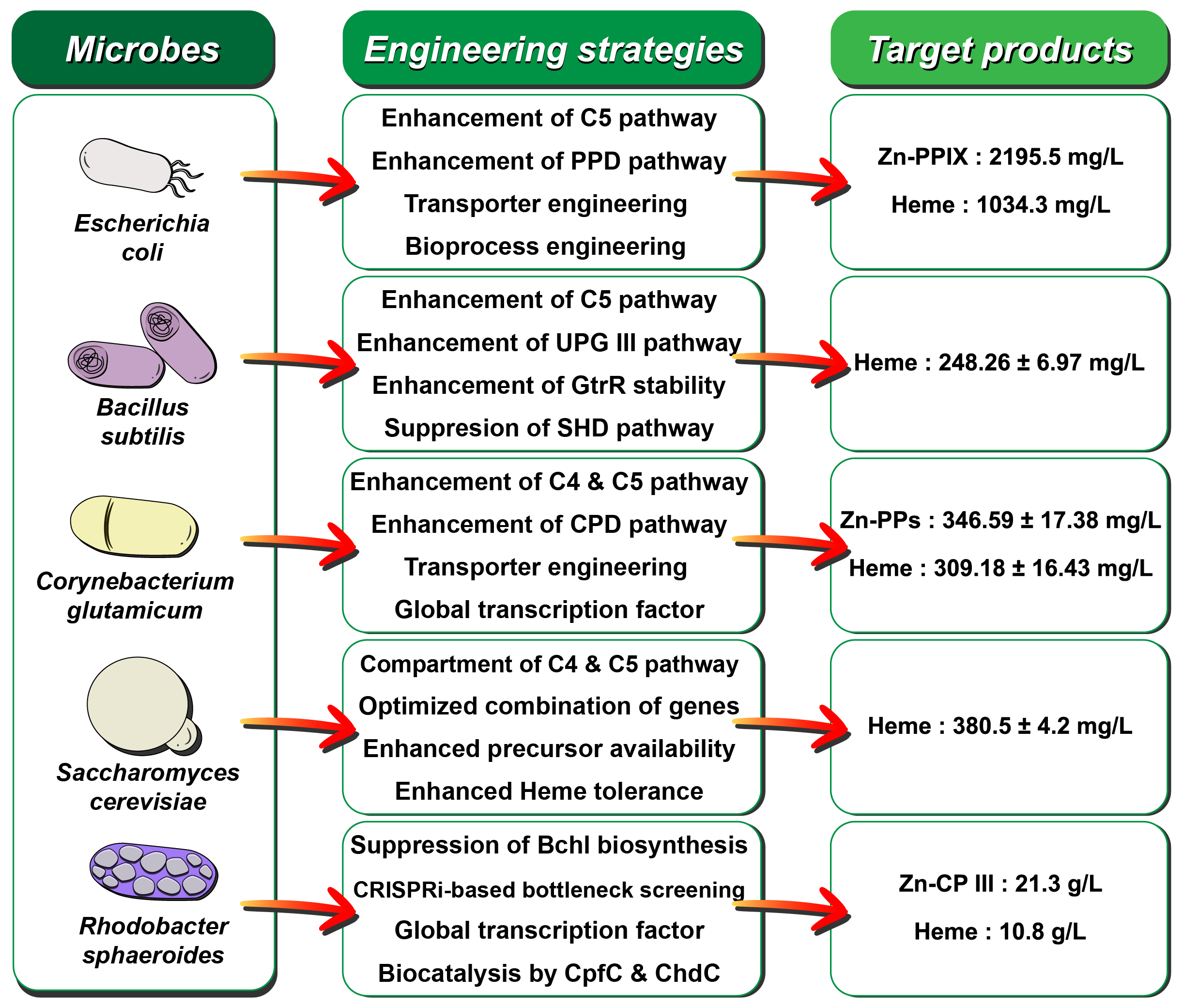
Fig. 1.
The major microorganisms and metabolic engineering strategies studied for enhancing metalloporphyrin biosynthesis. Additional information was referenced from (Ko et al. 2024).
Porphyrin Biosynthesis Pathways
The common core pathway for the formation of tetrapyrrole structures
Biosynthesis of metalloporphyrins begins from aminolevulinic acid (ALA) that is a common precursor, which is produced via two distinct pathways (Fig. 2)(Dailey et al. 2017, Layer 2021). One is the C4 pathway that exists in fungi and bacteria within the Pseudomonadota phylum. In this pathway, aminolevulinate synthase (AlaS) catalyzes ALA biosynthesis by condensing two substrates, glycine and succinyl coenzyme A (CoA), directly with the release of CO2 and CoA (Nandi 1978). AlaS is pyridoxal-5’-phosphate (PLP)-dependent. The other is the C5 pathway present in bacteria and archaea, ALA biosynthesis of which begins from L-glutamate and proceeds via the following three proteins: First, glutamyl-tRNA synthetase (GltX) catalyzes the aminoacylation of L-glutamate and tRNAglu to form L-glutamyl-tRNAglu. In the subsequent reaction, NADPH-dependent glutamyl-tRNA reductase (GtrR), the dimeric enzyme, reduces L-glutamyl-tRNAglu to glutamate-1-semialdehyde (GSA) with tRNAglu release (Hoober et al. 1988, Luer et al. 2005). Finally, glutamate-1-semialdehyde aminotransferase (GsaM), a PLP-dependent dimeric enzyme, catalyzes the interconversion reaction of the amino group and carbonyl moiety of GSA intramolecularly to form ALA (Sorensen and Stetefeld 2011). Interestingly, it was revealed that GtrR and GsaM formed a complex for substrate channeling of solvent-sensitive and unstable GSA (Luer et al. 2005). Then, two moles of ALA are condensed into one mole of porphobilinogen (PBG), a monopyrrole, by porphobilinogen synthase (PbgS), which is the homo-octamer requiring divalent metals for catalytic activity (Breinig et al. 2003). Hydroxymethylbilane synthase (HmbS) catalyzes the reaction through the enzyme substrate complex that links four molecules of PBG together in a line to form one mole of hydroxymethylbilane (HMB), a linear form of tetrapyrrole, with the release of four moles of ammonia (Jordan and Berry 1981). Finally, uroporphyrinogen synthase (UroS) inverts the fourth monopyrrole of linear HMB to form a type III isomer and cyclizes it to generate uroporphyrinogen III (UPG III), which is the macrocylic tetrapyrrole and the common intermediate of multiple pathways for heme biosynthesis (Layer 2021).
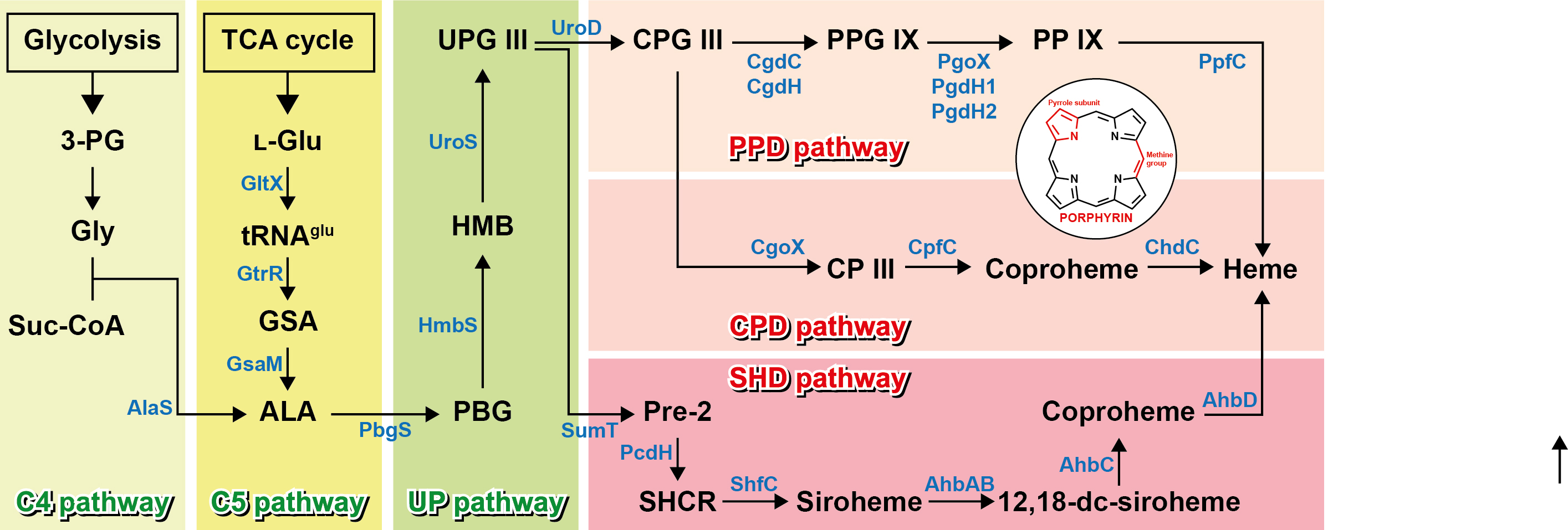
Fig. 2.
Metabolic pathways of porphyrins. Abbreviation: 3-PG, 3-phosphoglycerate; Gly, glycine; Suc-CoA, Succinyl-CoA; l-Glu, l -glutamate; GSA, glutamate-1-semi-aldehyde; ALA, 5-aminolevulinic acid; PBG, porphobilinogen; HMB, hydroxymethylbilane; UPG III, uroporphyrinogen III; CPG III, coproporphyrinogen III; PPG IX, protoporphyrinogen IX; PP IX, protoporphyrin IX; CP III, coproporphyrin III; Pre-2, Precorrin-2; SHCR, sirohydrochlorin; 12,18-dc-siroheme, 12,18-didecarboxy-siroheme; UP pathway, Uroporphyrin pathway.
Protoporphyrin-dependent pathway
The PPD pathway, the first discovered route for heme biosynthesis from UPG III, is recognized as the classical pathway. At first, the PPD pathway was thought to be the sole route to synthesizing heme in living organisms. However, the PPD pathway exists in some Gram-negative bacteria and eukaryotes, and other pathways, such as CPD and SHD pathways, have been discovered in Gram-positive bacteria, archaea, and sulfur-reducing bacteria (Dailey et al. 2017). Uroporphyrinogen III decarboxylase (UroD) catalyzes the stepwise decarboxylation of UPG III to yield coproporphyrinogen III (CPG III), which is the same reaction in the CPD pathway described later. UroD, a homodimer, converts four acetate groups of each pyrrole ring into methyl groups releasing four CO2 without the need for any cofactor. Then, CPG III is converted into protoporphyrinogen IX (PPG IX) via the decarboxylation reaction in which each propionic acid group on rings A and B of CPG III is converted to vinyl groups. There are two different mechanisms for the decarboxylation reaction of CPG III (Heinemann et al. 2008). The first is the oxygen-dependent decarboxylation reaction catalyzed by coproporphyrinogen III oxidase (CgdC), which is a monomer found in eukaryotes and some Gram-negative bacteria that requires molecular oxygen during the reaction and releases peroxide and CO2 at the end of the reaction (Breckau et al. 2003). The second is the oxygen-independent decarboxylation reaction catalyzed by coproporphyrinogen III dehydrogenase (CgdH), which is a homodimer found in other bacteria. This enzyme, which contains the [4Fe-4S] cluster, utilizes S-adenosylmethionine (SAM) molecules for the reaction and releases CO2 at the end of the reaction (Rand et al. 2010). After that, PPG IX is converted into protoporphyrin IX (PP IX) via a six-electron oxidation. There are three enzymes that catalyze the oxidation reaction (Layer 2021). The most studied enzyme is protoporphyrinogen IX oxidase (PgoX) found in some bacteria and eukaryotes. PgoX, the oxygen-dependent and flavin adenine dinucleotide (FAD)-containing homodimer, transfers electrons from PPG IX to the electron acceptor molecular oxygen with the aid of the cofactor FAD and then releases H2O2 molecules (Koch et al. 2004). On the other hand, most Gram-negative bacteria have one of the protoporphyrinogen IX dehydrogenases (PgdH1 and PgdH2), both of which are oxygen independent (Mobius et al. 2010). PgdH1 is a flavodoxin family protein containing flavin mononucleotide (FMN) as a cofactor and is found in γ-proteobacteria. It has been reported that this enzyme transfers electrons to the anaerobic respiratory chain using menadione or menaquinone as the final electron acceptor. Although much less is known about the biochemistry of PgdH2, this enzyme appears to utilize an anaerobic respiratory chain for electron transfer via a mechanism distinct from PgdH1 (Dailey et al. 2017). Finally, heme is biosynthesized via the chelation of ferrous iron to PP IX by protoporphyrin IX ferrochelatase (PpfC).
Coproporphyrin-dependent pathway
The CPD pathway is the most recently uncovered pathway which is mainly found in Gram-positive bacteria (Dailey et al. 2015). This route includes coproporphyrin III (CP III) as an intermediate but not PP IX. First, CPG III is converted into CP III with the release of three H2O2 molecules via six-electron oxidation catalyzed by coproporphyrinogen III oxidase (CgoX). CgoX, the FAD-containing protein, requires three O2 molecules as electron acceptors, and its mechanism is identical to that of PgoX (Lobo et al. 2015). It has been suggested that although CgoX also has a high degree of similarity to PgoX in the PPD pathway, there are significant differences in the size and charge of the active site pocket (Layer 2021). Next is the chelation of iron into CP III, which yields coproheme and is catalyzed by coproporphyrin III ferrochelatase (CpfC) found only in CPD pathway-possessing bacteria (Lobo et al. 2015). As mentioned above, even though the mechanism and structure of CpfC are very similar to those of PpfC, it has been predicted that CpfC has one less lip on the active site than PpfC. The lack of a lip might allow CP III that has two propionate groups on rings A and B, to enter the active site of CpfC (Layer 2021). Finally, heme is biosynthesized via the oxidative decarboxylation of coproheme by coproheme decarboxylase (ChdC). In this reaction, two propionate side groups chains on rings A and B are converted step by step into vinyl groups with the release of CO2 (Hofbauer et al. 2016).
Siroheme-dependent pathway
The oldest heme biosynthetic pathway from an evolutionary perspective is the SHD pathway using siroheme as a pivotal intermediate, which is present in sulfate-reducing bacteria and archaea (Ishida et al. 1998). Unlike the PPD pathway that converts UPG III into CPG III via decarboxylation in the first step, the SHD pathway converts UPG III into precorrin-2 via methylation. S-adenosyl-L-methionine uroporphyrinogen III methyltransferase (SumT) catalyzes the methylation of UPG III at the C-2 and C-7 positions to yield precorrin-2, which is a common precursor for the biosynthesis of siroheme, heme d1, vitamin B12, and coenzyme F430. Then, NAD+-dependent precorrin-2 dehydrogenase (PcdH) oxidizes the C-15 methine bridge of precorrin-2 to yield sirohydrochlorin (Storbeck et al. 2010). After that, siroheme is formed via the chelation of iron to sirohydrochlorin catalyzed by sirohydrochlorin ferrochelatase (ShfC) (Bali et al. 2014). The next step is the decarboxylation of siroheme at carbon positions 12 and 18 to produce 12,18-didecarboxysiroheme. Siroheme decarboxylases (AhbAB) are involved in this decarboxylation reaction (Palmer et al. 2014). Then, didecarboxysiroheme decarboxylase (AhbC) removes two acetic acid side chains at positions C-2 and C-7 in 12,18-didecarboxysiroheme to yield coproheme (Bali et al. 2014). Finally, heme is produced from coproheme via the decarboxylation of propionate chains on rings A and B to form vinyl groups by coproheme decarboxylase (AhbD).
Based on these porphyrin pathways, various studies have optimized metabolic pathways to enhance metalloporphyrin production. The following section reviews research findings on the improved production of metalloporphyrins using various microorganisms (Table 1).
Table 1.
Summary of metabolic engineering strategies for metalloporphyrin production and production profiles in various microorganisms
| Microbial producer | Engineering strategy | Production Titer (mg/L) | Ref. |
| B. subtilis |
1. Enhancing ALA Biosynthesis: Overexpression of GtrR with the knockout of the membrane protein HemX (which reduces GtrR concentration), ALA exporter YwfM, and glutamate dehydrogenase RocG (which converts glutamate to α-ketoglutarate); introduction of various AlaS enzymes. 2. Strengthening Downstream Pathways: Overexpression of PbgS, HmbS, and UroS (involved in UPG III biosynthesis) using the strong PlapS promoter; overexpression of SumT (involved in precorrin-2 biosynthesis in the SHD pathway); knockout of heme oxygenases HmoA and HmoB to prevent heme degradation. |
Heme: 248.26 ± 6.97 mg/L | Yang et al. 2023 |
| S. cerevisiae |
1. Enhancing ALA Biosynthesis: Overexpression of AlaS in mitochondria and GtrR and GsaM in the cytoplasm. 2. Strengthening Downstream Pathways: Overexpression of PbgS, HmbS, UroD, and CgdC; knockout of transcription factors HAP1 and ROX1 to mitigate the inhibitory effects of heme on CgdC expression. 3. Improving Tolerance to Heme Toxicity: Adaptive evolution using a CRISPR-assisted random mutation (CARM) approach to reduce heme cytotoxicity; overexpression of SPS22, PHO84, REE1, HEM4, and CLB2, which are involved in cell wall integrity, phosphate transport, metabolic regulation, and cell division. |
Heme: 380.5 mg/L | Guo et al. 2024 |
| R. sphaeroides |
1. Enhancing CP III Biosynthesis: Deletion of fnrL (a global anaerobic regulator controlling bacteriochlorophyll biosynthesis); knockout of cgdH based on genome-wide CRISPR interference (CRISPRi) screening to identify bottlenecks in CP III biosynthesis; mutation of the PrrAB two-component regulatory system, which senses redox states and regulates porphyrin metabolism. 2. Optimizing Metalloporphyrin Biosynthesis: Engineering of ferrochelatase (CpfC) and coproheme decarboxylase (ChdC) via mutagenesis to enhance heme and Zn-CP III production. |
Heme: 10.8 g/L Zn-CP III: 21.3 g/L | Chen et al. 2024 |
Recent Advances in Metabolic Engineering of Various Microorganisms for the Improved Production of Metalloporphyrins
Escherichia coli
E. coli was the first bacterium to perform metabolic engineering for the enhanced production of metalloporphyrins (Ko et al. 2024). One of the most recent achievements in E. coli carried out metabolic engineering and bioprocess optimization for heme production (Zhao et al. 2018). Zhao and colleagues successfully enhanced heme production by overexpressing six enzymes (GltX, GtrR, GsaM, PbgS, HmbS, and UroS) for ALA and UPG III biosynthesis, along with four key enzymes of the PPD pathway, using various vector combinations in E. coli. Additionally, they improved heme secretion in the engineered E. coli by overexpressing transporters and suppressing heme degradation and byproduct formation. Subsequently, this research group further improved heme production using the engineered strain by implementing bioprocess engineering strategies (Choi et al. 2022). Through optimization of pH conditions, iron and carbon source concentrations, and IPTG induction, a heme production yield of 1034.3 mg/L and a productivity of 21.5 mg/L/h were achieved.
Bacillus subtilis
A recent effort of metabolic engineering in B. subtilis has focused on improving heme production (Yang et al. 2023). This study enhanced heme biosynthesis by strengthening the ALA biosynthesis, UPG III biosynthesis, and downstream pathways. First, to improve the stability of the key enzyme GtrR in the C5 pathway, the membrane protein HemX, which reduces GtrR concentration, was knocked out. This modification increased heme concentration by approximately 2.9-fold. Additionally, the overexpression of GtrR with the knockout of the ALA exporter YwfM significantly improved both ALA and heme production. Furthermore, knocking out the rocG gene encoding glutamate dehydrogenase, which converts glutamate to α-ketoglutarate, further enhanced ALA and heme biosynthesis. Alongside the C5 pathway, introducing various AlaS enzymes from different origins into the C4 pathway slightly increased ALA biosynthesis. Next, the genes encoding PbgS, HmbS, and UroS, which are involved in UPG III biosynthesis, were overexpressed using the strong promoter PlapS, resulting in increased heme production. Finally, to strengthen the downstream pathway, SumT (referred to as NasF in the study) which participates in precorrin-2 biosynthesis in the SHD pathway as well as the heme oxygenases HmoA and HmoB which degrade heme were knocked out, resulting in enhanced heme production. In addition, fed-batch fermentation was performed over 144 hours using the final recombinant strain, achieving a heme production level of 248.26 ± 6.97 mg/L.
Corynebacterium glutamicum
C. glutamicum, known as a glutamate-overproducing strain, offers potential advantages for porphyrin biosynthesis. For this reason, it has recently been employed for the microbial production of both heme and Zinc-porphyrins (Zn-PPs). In a previous study for heme production, the integration of the C4 and C5 pathways successfully enhanced ALA biosynthesis and porphyrin pool (Ko et al. 2021). Notably, this research utilized the recently uncovered CPD pathway (including CgoX, CpfC, and ChdC enzymes) for the first time in metabolic engineering, achieving the further improvement in heme production. Additionally, a transcription factor DtxR associated with heme homeostasis was utilized to globally upregulate the porphyrin metabolic pathway, leading to enhanced heme biosynthesis. The study also discovered and optimized the binding of heme to cell wall and membrane proteins in Gram-positive bacteria, successfully improving heme secretion. Based on these metabolic engineering strategies, a strain capable of producing Zn-uroporphyrin III and Zn-coproporphyrin III was further developed, achieving a total Zn-PPs production of 346.59 ± 17.38 mg/L (Ko et al. 2023). Notably, Zn-PPs were purified to a purity exceeding 75% directly from the culture medium, and the purified compounds were successfully demonstrated as a potential sunscreen. Furthermore, they showed hydroxyl radical scavenging activity and displayed a synergistic effect with lignin, which indicates their potential as a bio-based sunscreen material.
Saccharomyces cerevisiae
Compared to other microorganisms, studies on heme biosynthesis via metabolic engineering in S. cerevisiae have emerged relatively recently. In a recent study, Lee and colleagues enhanced heme production in S. cerevisiae by optimizing the heme biosynthetic pathway, inhibiting heme degradation, and enhancing PP IX uptake (Lee et al. 2024). First, the researchers identified rate-limiting enzymes for heme biosynthesis by combinatorially overexpressing HEM2 (PbgS), HEM3 (HmbS), HEM12 (UroD), and HEM13 (CgdC). To improve precursor availability and prevent heme degradation, they overexpressed PUG1 (protoporphyrin uptake protein 1) to enhance PP IX uptake and disrupted HMX1 (heme oxygenase 1). These modifications resulted in significantly increased heme production.
Another recent study achieved higher heme production through multidimensional metabolic engineering of S. cerevisiae (Guo et al. 2024). Enhanced ALA synthesis was accomplished by overexpressing AlaS in the mitochondria (C4 pathway) and GtrR and GsaM in the cytoplasm (heterologous C5 pathway) by compartment. Similar to the earlier study, the overexpression of PbgS, HmbS, UroD, and CgdC was critical for increasing heme production. Furthermore, to address the inhibitory effects of heme on CgdC expression, the transcription factors HAP1 and ROX1 were deleted, which significantly improved heme production. Finally, to mitigate cytotoxicity of heme, a CRISPR-assisted random mutation (CARM) approach was employed for adaptive evolution, resulting in strains with improved tolerance to heme toxicity and reduced reactive oxygen species (ROS) levels. Heme production was further enhanced by overexpressing genes associated with specific cellular processes: SPS22 (encoding an ascospore wall assembly protein for cell walls), PHO84 (encoding a high-affinity inorganic phosphate (Pi) transporter for transmembrane transport), REE1 (encoding a regulator of enolase expression), HEM4 (encoding UroS), and CLB2 (encoding a B-type cyclin for cell division). Finally, a two-stage fed-batch fermentation process was implemented using the final engineered strain, separating biomass accumulation and heme production phases. This strategy achieved a high production yield of 380.5 mg/L with a productivity of 4.2 mg/L/h.
Rhodobacter sphaeroides
A recent study has demonstrated the potential of R. sphaeroides, a purple non-sulfur photosynthetic bacterium, as an efficient microbial chassis for metalloporphyrin production through metabolic engineering and biocatalysis (Chen et al. 2024). First, R. sphaeroides subspecies were engineered for CP III production by deleting of the fnrL gene, a global anaerobic regulator related to the regulation of bacteriochlorophyll (Bchl) biosynthesis. A fnrL-deficient R.sphaeroides showed a 6.2-fold increase in CP III production (480.2 mg/L) compared to the wild type, resulting in inhibiting downstream pathways involved in Bchl biosynthesis. Next, genome-wide CRISPR interference (CRISPRi) screening was performed to identify bottlenecks in CP III biosynthesis. The screening revealed that hemN (CgdH encoding oxygen-independent coproporphyrinogen oxidase) limited CP III accumulation. Deleting hemN in the engineered strain led to a significant increase in CP III production, reaching 1.25 g/L, representing a 160% improvement over the fnrL-deficient strain. Additional optimization focused on the PrrAB two-component regulatory system, which senses redox states and regulates porphyrin metabolism. By fine-tuning the phosphorylation of PrrA through a mutated histidine kinase (PrrBpLB2), CP III production was further increased to 16.5 g/L in fed-batch fermentation, demonstrating the potential for industrial-scale production. To convert CP III into metalloporphyrins such as heme and Zn-CP III, ferrochelatase (CpfC referred to as HemH in the study) from Staphylococcus aureus and coproheme decarboxylase (ChdC referred to as HemQ in the study) were used for biocatalysis and further engineered through random mutagenesis, resulting in a high-activity variant (HemQ28-M). Using a fusion enzyme system combining HemH and HemQ28-M, the process achieved a heme yield of 10.8 g/L and a Zn-CP III yield of 21.3 g/L in a 5-L bioreactor. This study provides a sustainable platform for high-yield production of metalloporphyrins, paving the way for broader industrial applications.
Conclusion
This study demonstrates the potential of metabolic engineering and biocatalysis to maximize metalloporphyrin production. By addressing pathway bottlenecks, cytotoxicity, and scalability, significant progress was achieved in enhancing production yields. These findings establish a foundation for the sustainable and industrial-scale synthesis of metalloporphyrins, enabling promising applications such as photodynamic therapy, photosensitizers, antibacterial agents, and functional foods across medical, industrial, and environmental sectors.
