

Introduction
Materials and Methods
Comammox maintaining
pH batch experiment
Chemical analysis
N2O measurements
Results and Discussion
Effect of pH levels on the activity of comammox N. inopinata with 1 mM NH4+ addition
pH effect on the NH2OH and N2O production from comammox
Conclusions
Introduction
Nitrification is a critical part of the global biogeochemical nitrogen (N) cycle. For more than a century, nitrification was recognized as a two-step process driven by two distinct microorganisms: (1) ammonia (NH3) oxidation to nitrite (NO2‒) by ammonia-oxidizing bacteria (AOB); (2) nitrite (NO2‒) oxidation to nitrate (NO3‒) by nitrite-oxidizing bacteria (NOB). The knowledge of nitrification has been updated largely in the past 20 years. The isolation of marine ammonia-oxidizing archaea (AOA) Nitrosopumilus maritimus in 2005 has inspired the studies on the cultivation of new AOA strains, physiology, and genomes of pure AOA strains and ecological niches of AOA in different ecosystems (Hatzenpichler et al. 2008, Könneke et al. 2005, Lehtovirta-Morley et al. 2011, Tourna et al. 2011). NOB were traditionally perceived as physiologically restricted organisms and were less intensively studied than other nitrogen-cycling microorganisms. However, new discoveries of an unexpectedly high diversity of mostly uncultured NOB and a great physiological versatility (Daebeler et al. 2020, Daims et al. 2016, Kitzinger et al. 2018, Koch et al. 2014). One intriguing finding was the discovery of a new microorganism in the genus of canonical NOB Nitrospira that can perform complete ammonia oxidation to nitrate (comammox) relatively recently (Daims et al. 2015, Van Kessel et al. 2015). In the majority of ecosystems, AOA, AOB, comammox, and NOB often co-occur. However, the factors that affect niche differentiation of the different groups remain unclear.
Many environmental and physiological factors are known to affect the niche differentiation and habitat selection of nitrifying microorganisms (Lehtovirta-Morley 2018). pH is an important driver for the community composition and adaptation (Gubry-Rangin et al. 2011, 2015). pH can affect the NH3 concentration in environments, with a significant reduction of NH3 below pH 7 as the pKa for the NH4+ to NH3 equilibrium is 9.25. Decreasing pH also moves the NO2‒ to HNO2 equilibrium toward inhibitory nitrous acid, which is highly reactive, with breakdown products that can cause extensive cellular damage. In addition, growth at low and high pH requires mechanisms for pH homeostasis to maintain the transmembrane proton gradient required for ATP production and normal function of cellular processes (Herbold et al. 2017, Wang et al. 2019). Most of the reported AOA and AOB grow at pH 7–8, with a slight difference in pH range among the different organisms. Certain nitrifiers can stand low or high pH and separate their ecological niche from other nitrifiers. For instance, Candidatus Nitrosotalea devanaterra (one soil AOA isolate) is the only soil AOA that can grow in the pH range of 3–5.5 (Lehtovirta-Morley et al. 2011), while Ca. Nitrosoglobus terrae is the only purified AOB that can grow at low pH (5–7.5) (Hayatsu et al. 2017). Recent research demonstrates that one Nitrospira species can grow at hapoalkali and halosanine conditions (Daebeler et al. 2020), indicating the potential for Nitrospira to withstand high pH. However, the pH range of comammox has not been reported. It would be interesting to know if the NO2‒ oxidation could help comammox oxidize NH3 at low pH.
Nitrification, especially ammonia oxidation, is an important source of nitrous oxide (N2O) production from terrestrial environments. AOB are recognized as a major source of N2O production from terrestrial environments (Gödde and Conrad 1999), and AOA are also considered to be important contributors to N2O production in various environments, based on their high abundance in many ecosystems and the documented formation of N2O during AOA-mediated ammonia oxidation (Jung et al. 2014, Kozlowski et al. 2016, Stieglmeier et al. 2014). In neutral pH conditions, the produced N2O from comammox is similar to that from AOA, but significantly lower than AOB (Kits et al. 2019). Comammox encodes ammonia oxidation machinery that is more closely related to AOB than to AOA. However, comammox does not encode nitric oxide reductase (NOR), which is similar to that of AOA. Isotope labeling experiments suggested hybrid N2O formation in the AOA Nitrososphaera viennensis from nitrite and an intermediate of ammonia oxidation, and was attributed to either an enzymatically catalyzed (codenitrification) or abiotic N-nitrosation reaction (Kozlowski et al. 2016). Furthermore, it was recently demonstrated that N2O is formed abiotically under aerobic conditions from hydroxylamine (NH2OH) and nitrite produced by aerobic ammonia-oxidizing microbes and that this hybrid pathway can account for a large proportion of the aerobically produced N2O (Liu et al. 2017). Most of the studies of N2O production by AOA and AOB have been performed in circumneutral pH ranges only, although various biological and chemical reactions involved in N2O production are pH-dependent. It has been reported that more N2O may be formed from AOA and AOB at lower pH compared to that at neutral pH (Jung et al. 2019), which could be due to a novel enzymatic reaction related to the enzyme cytochromes P450 that may function as a NO reduction pathway at lower pH. Like AOA, comammox owns a nitrite reductase gene (nirK) but lacks canonical NOR genes. Even though comammox seems closer to AOB based on the phylogenetic tree of AMO and HAO, it has quite similar traits of AMO affinity (Km) and N2O release rates with those of AOA (Kits et al. 2017, 2019). The NH2OH release rate of comammox was quite similar to that of AOB (Liu et al. 2017). Researches found that more N2O was produced at lower pH than that at neutral pH for both AOA and AOB (Jung et al. 2019). Interestingly, comammox also contains cytochrome P450; it is still unknown if the cytochrome P450 in comammox would activate at lower pH or not.
Therefore, the main goals of this study are: (1) to summarize the pH range of the reported AOA, AOB, and NOB, and compare the pH range with that of comammox; (2) to study the N2O and NH2OH production of comammox at low pH. We hypothesize that comammox may have a different pH range compared to that of canonical AOB, AOA, and NOB; and comammox may have the potential to produce more N2O at lower pH, just like AOA and AOB.
Materials and Methods
Comammox maintaining
Cultures of Nitrospira inopinata were maintained at 37°C with 1 mM NH4Cl in a CaCO3-buffered AOM medium containing (per litre) (Lebedeva et al. 2011): 50 mg KH2PO4, 50 mg MgSO4 × 7H2O, 75 mg KCl, 584 mg NaCl, 4 g CaCO3 (solid buffer), 1 ml selenium-wolframate solution (SWS), and 1 ml trace element solution (TES). For a detailed description of the composition of TES and SWS, please refer to Widdel (Widdel 1980). The pH of the medium was around 7.8–8.2 due to the CaCO3 buffer system during the culture growth.
pH batch experiment
Before the experiment, the stock cultures were transferred twice from CaCO3-buffered medium to 2 mM NaHCO3- buffered medium to remove the CaCO3 solids in the culture. Then, metabolically active cultures (200 ml) were harvested by centrifugation (8000 g × 15 min), washed, and resuspended in 10 ml 2 mM NaHCO3 buffered medium for preparing the inoculum. Buffers of 10 mM MES (2-(N-morpholino) ethanesulfonic acid), HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid), and TAPS ([tris(hydroxymethyl)methylamino] propanesulfonic acid) were used for adjustment of pH. The MES stock buffer was prepared by dissolving 9.76 g MES in 100 mL of 136 mM NaOH. The HEPS buffer stock was prepared by dissolving 23.83 g HEPS in 100 mL 600 M NaOH. The TAPS buffer stock was prepared by dissolving 24.3 g TAPS in 100 mL 570 mM NaOH. Buffer stock solutions were diluted to 10 mM MES, HEPS, and TAPS. Then, sterilized 1 M HCl and 1 M NaOH solutions were added to the medium to adjust pH to 5.5, 6, 7, 8, 8.5, and 9, respectively. Subsamples of 1 ml were taken from the cultures after 0, 0.3, 1.3, 2.3, 3.3, 4.3, 5.3, 6.5, 8.5, 12, and 14 days for the different pH treatments. The subsamples were stored at –20°C immediately after sampling for further chemical analysis.
Chemical analysis
Inorganic N concentrations were measured by protocols published by (Hood-Nowotny et al. 2010). Combined NH3/NH4+ concentrations in culture solutions were determined by the indophenol blue method. NO2‒ concentrations were spectrophotometrically measured by the Griess reaction after reacting with sulfanilamide and N-1-naphthyl-ethylenediamine dihydrochloride. NO3‒ concentrations were measured using Vanadium chloride reduction to NO2‒ followed by the Griess reaction.
N2O measurements
Comammox cultures were monitored daily and harvested immediately once all the substrate was consumed (normally ~7–9 days) by centrifugation using 10 kDa-cutoff, Ultra-15 Centrifuge Filter units (Amicon, Darmstadt, Germany). About 500 mL of mid-exponential phase culture were harvested per replicate experiment for N. inopinata. Harvested biomass was washed twice with substrate-free medium and then resuspended in 10 mL of the same substrate-free medium. The biomass was then transferred into a 10 mL, double-port MR chamber (allowing no headspace) that was fitted with two MR injection lids and two glass coated stir bars. All MR experiments were performed in a recirculating water-bath at 37°C. O2 uptake was measured using a OX-MR oxygen microsensor (Unisense, Aarhus, Denmark). N2O concentrations were measured using an N2O-MR sensor (Unisense). Substrate (NH4+) was injected into the chamber via an injection port using either a 10-µL or 50-µL syringe (Hamilton, Reno, USA) fitted with a 26G needle. NH4+ injections into the 10 mL MR chamber were always 1 mM. The OX-MR and N2O-MR sensors were plugged directly into a microsensor multimeter. All electrodes were polarized for >1 day prior to use and calibrated according to the manufacture’s instructions. All data were logged on a laptop via the microsensor multimeter using SensorTrace Logger software (Unisense). The output from One-Channel Free Radical Analyzer was run into the microsensor multimeter using a BNC/lemo adapter.
Results and Discussion
Effect of pH levels on the activity of comammox N. inopinata with 1 mM NH4+ addition
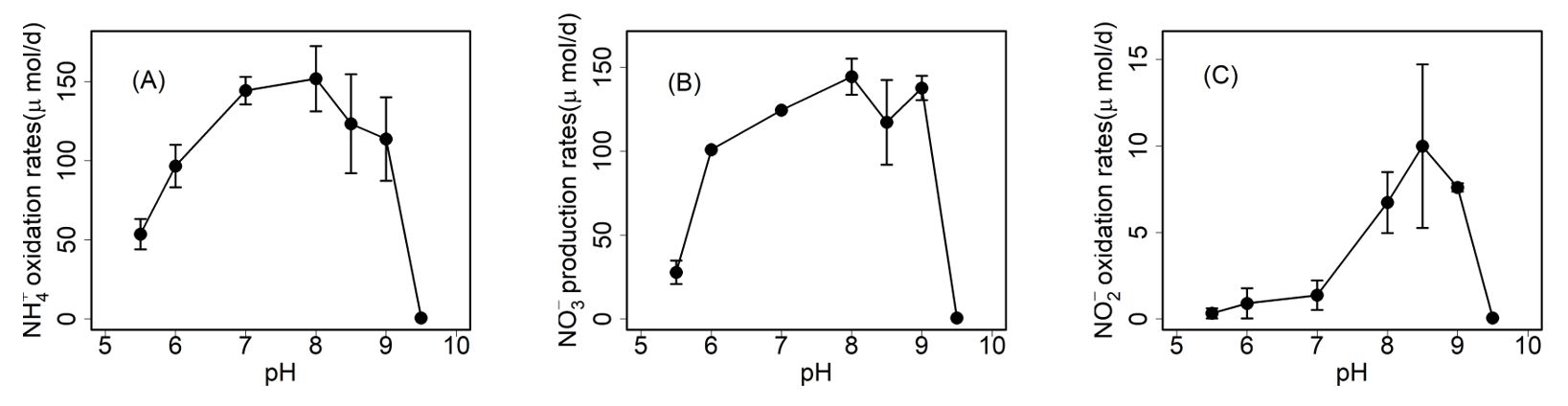
Comammox have ammonia oxidation activities in the pH range of 5.5–9. It can oxidize 1 mM NH4+ to NO3‒ completely in the pH range of 6.5–9, while it oxidizes only around 20% and 80% of added NH4+ at pH 5.5 and 6, respectively (Fig. 1A). The pattern of NO2‒ oxidation is similar with that of NH4+ oxidation, with least accumulation of NO2‒ at lower pH (5.5 and 6.0) compared to that at higher pH (8.5 and 9) (Fig. 1B). It appears that the oxidation of NO2‒ ceased at pH 5.5 in the later incubation period, which is consistent with the pattern of NH4+ oxidation at pH 5.5. From the reported pH range of other canonical AOB, AOA, and NOB, comammox seems to have a similar pH range to that of N. viennensis (Fig. 2). Generally, the pH range of AOA is narrower than that of AOB. AOA has activities in the pH range of 6.0–8.0, except N. devanaterra, which had activities in the pH range of 4–5.5. AOB has activities mostly in the pH range of 5.5–9.5, indicating the adaptation of AOB in different environments. AOB is more adapted to environments with higher pH, while AOA is more adapted to neutral or lower pH. For the canonical NOB, most of them have activities in the range of 6.0–8.0, except one acid Nitrobacter and one alkali Nitrospira. From the perspective of NH4+ oxidation rates, comammox has the largest NH4+ oxidation rates, NO3‒ production rates, and NO2‒ oxidation rates at around pH 8, 8–9, and 8.5, respectively, indicating that comammox prefers neutral and a little alkali (Fig. 3). Different nitrifiers have evolved diverse strategies to cope with pH stress. Some AOB and NOB form aggregates to maintain higher internal pH (De Boer et al. 1991), while certain nitrifiers (including AOB, AOA, comammox, and NOB) may have the genes to use urea, when NH3 concentrations are too low due to the low pH in surrounding environment (Burton and Prosser 2001, de Boer and Laanbroek 1989). Comammox and AOA are generally more tolerant of low pH due to their high affinity for NH3. For example, the acidophilic AOA Candidatus Nitrosotalea devanaterra grows at pH 3–5.5 (Lehtovirta-Morley et al. 2011), and molecular analyses have shown AOA dominance in acidic soils (Nicol et al. 2008, Zhang et al. 2012). Nonetheless, AOB can also prevail under acidic conditions; Candidatus Nitrosoglobus terrae grows at pH 5.5–7.0 (Hayatsu et al. 2017).

Fig. 1.
The effect of pH on the activity of Nitrospira inopinata with 1 mM NH4+ addition. Different pH levels are indicated as hollow circle (pH 5.5), hollow triangle (pH 6.0), solid rectangle (pH 7.0), solid circle (pH 8.0), solid triangle (pH 8.5), and solid diamond (pH 9.0). The values are presented as mean ± standard error (SD).
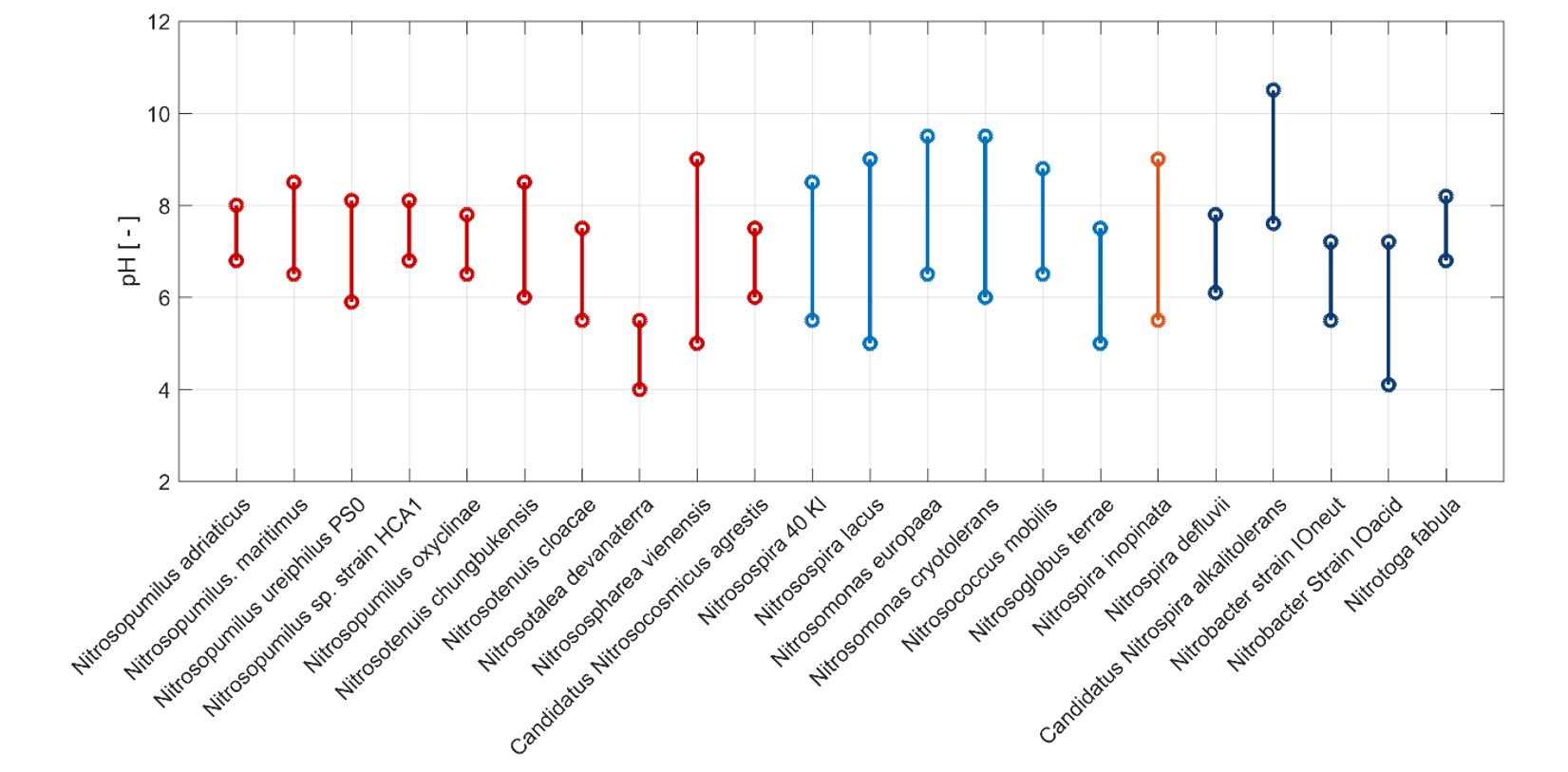
For NOB, most acid-tolerant isolates belong to the genus Nitrobacter, such as Nitrobacter sp. Io acid from forest soils (Hankinson and Schmidt 1988). In contrast, Nitrospira species are generally sensitive to low pH, likely due to high susceptibility to nitrous acid (Wang et al. 2016). Nevertheless, comammox populations have been reported as dominant in acidic soils (pH 4–7) (Hu and He 2017). In our study, N. inopinata exhibited a relatively low pH limit of 5.5, similar to some AOB. At the opposite extreme, comammox maintained full activity up to pH 9.0, comparable to AOB, and even alkali-tolerant Nitrospira have been reported to function up to pH 10 (Daebeler et al. 2020). These findings indicate that comammox Nitrospira possess broad pH adaptability, spanning both acidic and alkaline environments.
pH effect on the NH2OH and N2O production from comammox
Using NH3 as the electron donor, we measured instantaneous N2O production with microsensors by N. inopinata biomass at two pH levels (7.5 and 5.65) (Fig. 4). In contrast to the findings by Kits et al. (2019), N. inopinata produced N2O during aerobic NH3 oxidation and kept flat during anoxic period at pH 7.5. This could be due to that we added 1 mM NH4+ in the beginning and they only added 250 µM NH4+ in the beginning.
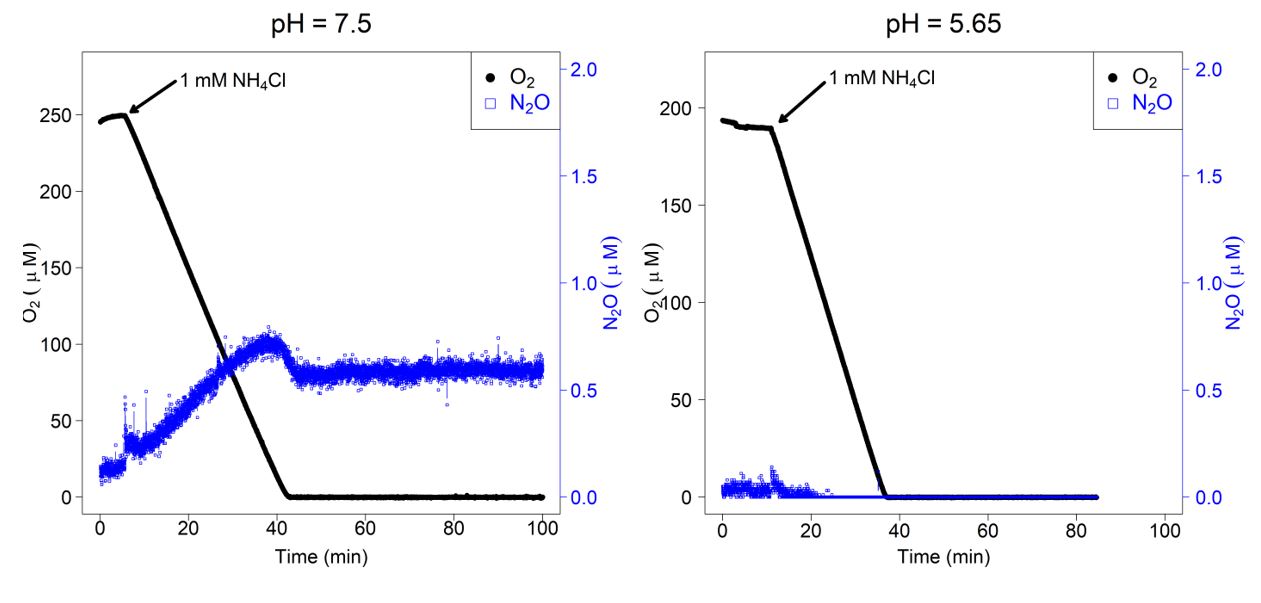
Fig. 4.
Instantaneous O2 consumption and N2O production during NH3 oxidation by N. inopinata. Dissolved O2 is shown in solid black circles, dissolved N2O in filled blue rectangles. Experiments were performed in a 10-mL microrespiration (MR) chamber fitted with O2 and N2O microsensors. The arrow marks the addition of 1 mM NH4Cl into the MR chamber.
AOB and AOA release small amounts of NH2OH during ammonia oxidation under fully oxic conditions and the abiotic conversion of NH2OH with Fe3+, Mn4+, Cu2+, NO2‒, and other components of the surrounding matrix under oxic conditions explains most of the N2O formation by these nitrifiers under these conditions (Liu et al. 2017). Decreasing pH leads to a significant increase in the N2O yield from NH3 in AOB and AOA (Jung et al. 2019), while the pH effect on the N2O yield in comammox remains unclear. Main pathways that contribute to N2O formation during acidification in AOB are nitrifier-denitrification, the sequential enzymatic reduction of NO2‒ to N2O via NIR and NOR, and chemodenitrification, whereby NO2‒ or NO is non-enzymatically reduced to N2O via media components or heat-killed cell moieties. AOA cannot perform nitrifier-denitrification to N2O due to the lack of NOR, also releases large quantities of NO during hypoxic conditions, which then reacts abiotically with medium components (Cu2+ and Fe2+) to form N2O (Kozlowski et al. 2016). Interestingly, decreasing pH leads to a significant increase in N2O production in AOA. Changes of 15N site preference (SP) values within the N2O molecule (as indicators of pathways for N2O formation), caused by decreasing pH, were highly different between the tested AOA and AOB. While acidification decreased the SP value in the AOB strain, SP values increased to a maximum value of 29‰ in one AOA strain, N. oleophilus. In addition, 15N-nitrite tracer experiments showed that acidification boosted nitrite transformation into N2O in all strains, but the incorporation rate was different for each ammonia oxidizer. Unexpectedly, for N. oleophilus, more than 50% of the N2O produced at pH 5.5 had both nitrogen atoms from nitrite. The high SP value and high portion of N2O production from NO2‒ suggests that most of the N2O produced was from either abiotic reactions between nitrification intermediates, e.g. NH2OH and NO (Heil et al. 2014), or from fungal denitrification that was catalyzed by an enzyme cytochrome P450 as NOR (P450nor) (Shoun et al. 2012). Putative cytochrome P450-encoding genes were detected in the genomes of N. oleophilus, and subsequent experiments showed the transcription of these genes at lower pH conditions, indicating that N. oleophilus can produce N2O via fungal denitrification at lower pH. Interestingly, putative cytochrome P450-encoding genes were also detected in other nitrifying bacterial strains, including comammox Nitrospira inopinata (Jung et al. 2019). Therefore, it would be interesting to know if acidification would stimulate N2O production in comammox.
Conclusions
In this study, we demonstrated that Nitrospira inopinata exhibits a broad pH tolerance ranging from 5.5 to 9.0, with optimal ammonia and nitrite oxidation activities occurring near neutral to slightly alkaline conditions. While comammox tolerated low pH, its ammonia oxidation capacity was significantly reduced at pH 5.5, and N₂O production did not increase under acidic conditions—unlike in AOA and AOB. Our findings suggest that comammox may rely less on acid-induced enzymatic or abiotic pathways for N2O formation. The presence of putative cytochrome P450 genes in the genome of N. inopinata raises questions about their functional role under acidic conditions, which remains to be explored. Overall, this study broadens our understanding of the ecophysiological response of comammox to pH, and highlights the need for further investigations into its regulatory mechanisms and contribution to N2O emissions under environmentally relevant pH conditions.
